Hugo Fjelsted Alrøe
AT TÆNKE SIG EN TILBLIVELSE
Overvejelser om Forklaring og Forståelse
af Natur og Evolution.
|
"Guderne åbenbarede ikke fra begyndelsen alt for os, men i tidens løb kan vi gennem vor søgen vinde bedre erkendelse. Men den absolutte sandhed har intet menneske kendt. Ej heller vil han komme til det - hverken om guderne eller om alle de øvrige ting jeg beretter om. For selv om han tilfældigt ytrede den endelige sandhed, ville han ikke selv vide det: thi alt er kun et spind af gisninger." Xenofanes fra Kolofon, 570-475 f.Kr..
Hovedopgave ved lektor i videnskabsteori Carsten Bengt-Pedersen, på Den Kongelige Veterinær- og Landbohøjskole:
Af Hugo Fjelsted Alrøe Studienummer Hg 1377, KVL
Amager d. 24/3 1989 [HTML udgave, 8.mar.1998]
KAPITEL 1: INDLEDENDE BEMÆRKNINGER OM VIDENSKAB OG HISTORIE
1.1 Om er og bør
1.2 Om reduktioner
1.3 Tro og åbenhed
1.4 Om at bevise og modbevise
1.5 Videnskabelig metode
1.6 Bateson, natur og ånd
1.7 Videnskab, historie og myte
1.8 Historie og objektivitet
KAPITEL 2: INFORMATION OG KOMMUNIKATION
2.1 Kommunikation og sprog
2.2 Det emiske aspekt
2.3 Lidt om sansning
2.4 Et slag på kinden
2.5 Tilfældighed og valg
2.6 Kvantemekanik og biologi
2.7 Den virkelige verden
2.8 Opdelinger og grænser
2.9 Beskrivelse, forklaring og forståelse
2.10 Noget om sansning, genkendelse og forståelse
KAPITEL 3: EVOLUTION
3.1 Neodarwinisme
3.2 Artsbegrebet
3.3 Flere slags arter
3.4 Individets udvikling
3.5 Om at ekstrapolere
KAPITEL 4: DET HIERARKISKE ASPEKT
4.1 Systemer og hierarki
4.2 Logiske typer, klasser og paradokser
4.3 Logiske typer og kommunikation
4.4 Kommunikation og kontekst
4.5 Genetik og regulering
4.6 Regulering i ontogenesen
4.7 Nogle typer af proces eller udvikling
KAPITEL 5: INDIVID OG POPULATION
5.1 Forandring i individ og population
5.2 Arvelighed og reproduktion
5.3 Kønnet formering og meiosens betydning
5.4 Somatiske forandringer og adaptation
5.5 Genotypiske forandringer og tilpasning
5.6 Logiske niveauer
5.7 Evnen til at skelne
KAPITEL 6: AUTOPOIESE
6.1 Erkendelsens cirkulære natur
6.2 Selvopretholdelse
6.3 Information på ny
6.4 Autopoiesens arv
6.5 Strukturel kobling
6.6 Metacellulære enheder
6.7 Levende væseners naturlige drift
6.8 Teleologi og tilpasning
6.9 Adfærd og viden
6.10 Sociale fænomener
KAPITEL 7: DISSIPATIVE STRUKTURER
7.1 Tidens symmetri og irreversibilitet
7.2 Orden gennem fluktuation
7.3 Et hierarki af systemaspekter
7.4 Fluktuation og stabilitet
7.5 Symmetribrud og indeterminisme
7.6 Spænding og stabilitet
7.8 Information; nyhed og bekræftelse
KAPITEL 8: DEN HISTORISKE EVOLUTION
8.1 Darwin og historiske processer
8.2 Bricolage
8.3 Tilfældighedens rolle
8.4 Tilfældighedens rolle i et større perspektiv
8.5 Kladistisk systematisering af mønstre
8.6 Kladistisk analyse og fylogenier
8.7 Slags og slægtskab
KAPITEL 9: EVOLUTIONENS MEKANISMER
9.1 Kontekst og evolution
9.2 Forandring og konstans
9.3 Fyletisk gradualisme eller ej
9.4 Punctuated equilibria
9.5 Stabilitetens forklaringer
9.6 Homøostatisk stabilitet
9.7 Splittelse og social isolation
9.8 Opsplitning og isolationsbarrierer
9.9 Mulighedernes spil
9.10 Stokastiske processer i flere tidshorisonter
FORORD Indhold
Denne opgave må ses mest som et forsøg for mig personligt på at blive mere afklaret omkring et emne, et forklaringsgrundlag i mit liv.
Et forsøg på bedre at forstå hvad jeg er, hvor jeg hører til; et spejl at spejle min opfattelse af mig selv i.
Det er også min ambition at videregive noget som nemt forsvinder i vores betagelse over, hvad vi kan gøre med vores indsigt og teknik. Jeg vil forsøge at gøre opmærksom på det vi ikke ved, det vi blot kan fascineres af, og det vi i vores naivitet tror er simpelt.
Jeg håber, at læseren kan fornemme og forstå den ydmyghed jeg af og til føler; for ydmyghed er nødvendig, hvis vi skal overleve.
Endelig er denne opgave et forsøg på at lære andre og 'nye' måder at tænke om verden på, nye for mig og måske også for læseren, for derigennem at opnå en bedre forståelse; og dette er faktisk det vigtigste af det hele.
Nogle vigtige elementer i disse nye tænkemåder er historiens vigtighed, ubestemthedens rolle, reduktionernes begrænsninger og helhedernes dynamik.
Denne opgave har også karakter af et forsøg. Opgaven inddrager en mængde emner fra mange områder i vores verden; det er det generelle og samlende der er i fokus. Som følge heraf er de specielle områder hver for sig kun omtalt på en ret overfladisk eller kortfattet måde; dette skyldes på ingen måde en nedvurdering af det specielle.
Forsøget grunder i et ønske om overblik over og forståelse af større sammenhænge i vores verden.
Jeg føler tydeligt min begrænsning overfor opgaven; - men jeg tror det er nødvendigt at prøve.
Verden vokser i et skræmmende tempo; en vækst af viden og tekniske muligheder. Millioner af mennesker arbejder med at skabe vores verden, hver inden for deres lille område. Det er egentlig ikke selve væksten, der skræmmer mig. Det er snarere mangelen på ønske om og vilje og midler til at skabe sammenhæng i denne vækst og forståelse af, hvordan den verden der er og den verden vi skaber fungerer sammen.
For det systematiske blik er der dele der mangler, og dele der kunne undværes i opgaven. Systematikken er nyttig til at skabe overblik, men den må ikke være en betingelse. Systematik og nyskabelse står nemlig i et vist modsætningsforhold, og 'side'-spor kan ofte skabe nye forbindelser indenfor emnet og til emnets omverden.
Jeg har i opgaven forsøgt at give kreativiteten gode betingelser, og det er derfor ret vilkårligt i forhold til emnet, hvor stor udstrækning og vægt de enkelte kapitler og afsnit har fået, og hvilken rækkefølge de kommer i.
Jeg prøver at overholde en vis sparsomhed med ord og begreber, og jeg søger at forklare de nødvendige begreber efterhånden som de anvendes.
Jeg håber at videregive nogle tanker omkring vanskelige problemer, og hvis ikke læseren forstår mit sprog og mine begreber (mine redskaber) ser jeg ingen mulighed for, at hun kan forstå hvad jeg mener.
Jeg vil ikke i opgaven spørge om eller forsøge at svare på, hvem der har ret og hvem der ikke har, sådanne spørgsmål har ikke min interesse. Jeg vil prøve at bruge de tanker, jeg mener at se hos de forfattere jeg anvender, når jeg føler de bidrager til min forståelse.
Forståelse er ikke en universelt gangbar mønt; min forståelse er ikke forfatternes forståelse, og den vil heller ikke være læserens forståelse.
Jeg bruger citater i to situationer. Dels når jeg ikke føler at kunne udtrykke min forståelse af et emne på en bedre måde. Og dels når jeg vil bruge et synspunkt i argumentationen, som jeg ikke er enig i.
Men oftest omskriver og samskriver jeg den eller de tekster jeg arbejder udfra, og henviser til kilderne.
I nogle tilfælde har jeg fordrejet mening og indhold i så høj grad i forhold til kilden, at jeg undlader at henvise, eller skriver 'inspireret af', efter eget skøn.
Direkte citater må ikke opfattes som 'de citeredes opfattelse', de vil altid være taget ud af én sammenhæng og anbragt i en anden; det er den forståelse uddraget har givet mig i 'min' sammenhæng, jeg forsøger at give videre.
Jeg gentager visse opfattelser eller billeder ret ofte i opgaven, oftest i varierende tankerækker og sammenhænge. Dette er dels for hele tiden at holde antagelser og forudsætninger klart; men også for at knytte de forbindelser, der gerne skulle skabe et sammenhængende mønster eller en gennemgående forståelse i opgaven.
Det er mit håb at dette mønster skinner igennem og, at det kan inspirere andre som det har inspireret mig; men jeg håber samtidigt, at ingen vil opfatte noget i denne opgave som rigtigt i nogen endegyldig forstand.
Jeg vil afslutte forordet med et af de citater, der ikke skal omskrives. Fysikeren David Bohm skriver i 'Helhed og den indfoldede orden' om videnstilblivelse:
"... den tanke, hvis indhold er helheden, [må] opfattes som en kunstform, som poesi, hvis funktion primært er at fremkalde en ny erkendelse og handlinger i overensstemmelse hermed, snarere end at kommunikere en refleksiv viden om, 'hvordan alting er'. Dette indebærer, at der er lige så lidt mulighed for en 'endelig' tanke, som der er mulighed for et endegyldigt digt (som ville gøre alle efterfølgende digte overflødige).
Enhver tænkning om helheden kan tages som udtryk for en anskuelse af vores kontakt med virkeligheden. Men alle anskuelser er dog begrænsede i den betydning, at de kun inden for visse grænser kan føre til en samlet orden og harmoni; uden for grænserne ophører de med at være relevante og passende. I sidste instans skal tankebevægelsen, som omfatter enhver helhedsopfattelse, ses som en proces med konstant forandrende form og indhold. Hvis denne proces udføres på rette måde med opmærksomhed henledt på en klar forståelse af tanken i dens tilblivelsesstrøm, så vil man ikke få for vane stiltiende at behandle indholdet som en endelig og i alt væsentligt statisk virkelighed, der ville være uafhængig af tanken. ...
Det er i denne sammenhæng vigtigt at understrege, at hvis man til stadighed identificerer bestemte synspunkter vedrørende helheden med bestemte personer (f.eks. Whitehead), så kommer man i konflikt med en modsigelsesfri behandling af viden som en integreret del af en samlet proces. Hvem der end overtager Whiteheads synspunkter, benytter vedkommende faktisk disse som et udgangspunkt for en forsat proces af videnstilblivelse. (Vi kunne måske sige, at vedkommende arbejder sig videre op af 'kundskabens flod'.) I denne proces kan nogle aspekter godt forandre sig langsomt, hvorimod andre forandrer sig hurtigere, men den hovedpointe, man skal holde sig for øje, er, at processen ikke har noget definerligt aspekt, som ligger absolut fast. Man må benytte sig af en trinvis intelligent erkendelse for at opdage de aspekter, som bør forandre sig langsomt og de, som bør forandre sig hurtigt, imens man på kunstnervis arbejder med at skabe ideer om 'det værendes helhed'. ...
Hovedspørgsmålet er så: "Kan vi være bevidste om denne vidensproces' stadigt forandrende og strømmende virkelighed?"
Hvis vi kan tænke ud fra en sådan klar bevidsthed, vil vi ikke komme til fejlagtigt at antage det, som stammer fra tanken, for det, som stammer fra en virkelighed, som er uafhængig af tanken. Og således kan tænkning med helheden som indhold udvikle sig på en måde, som er fri for den forvirring, der er indbygget i de tankeformer, som én gang for alle prøver at definere, hvad 'hele virkeligheden er', og som derfor får os til at forveksle en sådan tankes indhold med den samlede orden i en total virkelighed, som er uafhængig af tanken."
(Bohm 86, s 81ff)
Amager, marts 1989
Hugo Fjelsted Alrøe.
Kapitel 1 Indhold
INDLEDENDE BEMÆRKNINGER OM VIDENSKAB OG HISTORIE
1.1 Om er og bør:
Er det rigtigt at slutte fra, hvordan naturen er til, hvordan vi bør handle?
Denne opfattelse kaldes 'den naturalistiske fejltagelse'. Det almindelige misbrug af begreberne natur og naturlig viser faldgruberne i en direkte slutning fra er til bør.
Vores fastlæggelse af hvordan naturen hænger sammen er jo som alt andet uhjælpeligt forankret i den kultur og de omgivelser, vi er vokset op i.
Men: "nok kan biologien ikke undslippe det vilkår, at dens konstruktion af naturen altid vil være socialt ladet, men et eller andet sted må denne konstruktion dog have med naturen selv at gøre!", siger Hoffmeyer, og jeg er enig med ham.
(Bl.a Hoffmeyer 84, s220f og Jacob 85, s 53ff)
Etik, moral og retfærdighed kan ikke begrundes i biologiske forhold, men de er heller ikke uden sammenhæng med naturvidenskaben. Ib Ravn skriver herom:
"... hvad sker der, hvis vi ... undlader at inddrage naturvidenskaben i vores overvejelser over hvordan livet, samfundet og verden hænger sammen?
Der sker dét der altid sker, når vi erklærer at det ikke er nødvendigt at tænke alt for meget om et givet emneområde: nemlig at det bliver de ubevidste og ureflekterede forestillinger, der kommer til at styre vores praksis og vores tænkning på området.
Disse forestillinger kommer fra det kulturelle arvegods, som vi alle bærer rundt på, det er de ting som alle véd og tager for givne. Sådanne ubevidste forestillinger er ofte mangelfulde. I dette tilfælde er de katastrofale."
(Ravn 88)
En bedre forståelse af hvordan verden er, er nødvendig for en 'rigtigere' beslutning om hvad vi bør gøre!
Jeg tror på, at videnskaben faktisk udvikler sig eller rettere, at den videnskabelige erkendelse (under visse former, men hvilke?) bevæger sig imod en bedre forståelse af verden; dette er nok en af mine grovere fordomme, og én jeg har det lidt ubehageligt med, men den er nødvendig for min sjælefred. Jeg mener, hvis vi ikke kan tro på muligheden for at opnå en bedre forståelse, hvad har vi så at hænge ansvarsfølelse, engagement og i sidste ende etik op på?
Der er vel ingen tvivl om, at de forudsigelser videnskaben vælger at fremsætte, bliver stadigt mere nøjagtige, men der er forskel på nøjagtighed eller viden om detaljer og en forståelse af hvordan vores verden hænger sammen. På den anden side er der for mig ingen modsætning i, at vi kan få en bedre og bedre forståelse uden nogensinde at komme tæt på en endegyldig sandhed. (Mange indenfor videnskaben troede faktisk før århundredskiftet, at man nu var tæt på at have den fulde sandhed om verden - bortset fra nogle småting.)
Vores mulige verden af erkendelse er ikke en lukket verden, jeg tror ikke der er nogen absolut grænse for nyskabelse i verden; erkendelse er også nyskabelse, og denne ring af skabelse og forståelse er uden ende.
1.2 Om reduktioner:
Erkendelse og forståelse af en uhyre kompleks verden som vores, må nødvendigvis bygge på reduktioner, hvis vi skal kunne videregive og dele forståelsen. Jeg benægter ikke, at erkendelse af komplekse sammenhænge kan være af intuitiv eller mystisk art og, at dette kan være et vigtigt og måske endda nødvendigt skridt mod voksende erkendelse. Men videregivelse af erkendelsen kan kun ske, hvis erkenderen er istand til at reducere den, - uddrage fragmenter af den, der er tilstrækkelig simple til at kunne kommunikeres.
Vi har ikke nogle effektive måder, hvorpå vi kan kommunikere om helheder, specielt er det videnskabelige sprog helt utilstrækkeligt hertil.
Hvis vi vil forstå naturen og den levende verden, er historien et af de aspekter der ikke må abstraheres fra, på trods af vanskelighederne ved at behandle den videnskabeligt.
Evolutionsteori er et forsøg på at forstå den evolutionære historie, og klarlægge dens processer og mekanismer. Opfattelsen af evolutionen spiller ind mange steder i den biologiske erkendelse, og den udgør et grundlag i det meste af den biologiske forskning. Fejlagtige opfattelser eller utilstrækkelig forståelse på dette område er derfor ensbetydende med, at vores viden og forståelse på andre, også mindre omdiskuterede, områder kan være opbygget på et utilstrækkeligt grundlag.
Studier af en organisme i sig selv giver ikke samme forståelse som studier af organismen-og-dens-historie eller organismen-i-dens-miljø-og-deres-historie.
Evolutionsteorien har en stor betydning i vore bestræbelser på at forstå sammenhænge i økosystemer og i (vores alle sammens) biosfæren.
Desuden spiller evolutionsteorien også ind i vores liv på en anden måde, den er en vigtig del af vores natursyn/menneskesyn i kraft af dens funktion som skabelsesmyte.
Men først må vi se lidt på videnskaben og dens metoder.
Jeg føler mig ikke særlig sikker på mine holdninger og meninger i almindelighed. Dette er ikke tilstrækkeligt til at undgå, at mine af min opvækst og kultur karakteriserede 'fordomme' ligger til grund for de her fremførte opfattelser.
'Fordomme', dvs. tro, antagelser eller grundholdninger vi ikke tager stilling til i de enkelte problemstillinger, må ikke bare ses som negative hindringer for øget forståelse, men også som et nødvendigt grundlag herfor - udsynet og nysynet må ske fra et ståsted.
Til gengæld er det evnen og viljen til at afprøve alternative ståsteder i løbet af livet, der forhindrer dogmatisme, åbner for forandring og udvikling, og bevirker den utryghed der er indeholdt i det at leve.
Denne holdning til kulturelle og personlige fordomme eller grundliggende holdninger er inspireret af naturhistorikeren og palæontologen Stephen Jay Gould, der igen refererer til Thomas Kuhn.
Kuhn udtrykker denne holdning omtrent således:
Viden om det forventede forsinker erkendelsen af det uventede. Men det uventede erkendes kun som nyt af den, der nøjagtig ved hvad der forventes (eller - uregelmæssighed kan kun ses på den baggrund der udgøres af paradigmet).
(Kuhn 73, s 92).
Paradigmer er fælles fordomme, holdninger, antagelser eller teorier indenfor videnskab og tænkning, der ligger uhyre fast, fordi resten af teoribygningen hviler på dem; de er næsten umulige at skifte ud.
Forskning indenfor et paradigme kalder Kuhn for normalvidenskab. Normalvidenskaben sætter ikke spørgsmålstegn ved paradigmets gyldighed. Iagttagelser og tankerækker, der strider mod paradigmet, bliver affærdiget som ubetydelige, irrelevante eller tilhørende en anden faglig disciplin, de bliver indpasset gennem specielle antagelser, gemt til senere løsning eller simpelthen ignoreret.
Efterhånden som disse 'uregelmæssigheder' bliver flere og alvorligere, bliver modsætningerne for store for nogle få forskere og disse erkender, at paradigmet selv er utilstrækkeligt, og arbejder på at formulere et nyt grundlag. Nogle af de andre forskere 'omvendes' til det nye paradigme, og nye forskere vil se en valgmulighed og (forhåbentlig) vælge det mest sammenhængende paradigme. Men især ældre forskere holder fast på det gamle paradigme hele deres liv.
Kuhn betragter konkurrerende paradigmer som helt forskellige, uforenelige og usammenlignelige (sic!), og normalvidenskab før og efter et paradigmeskift vil derfor ikke kunne vurderes eller sammenlignes i nogen objektiv forstand. Heri er jeg ikke enig.
(Kragh 81, s 169ff)
1.4 Om at bevise og modbevise:
Jeg har for nyligt stiftet bekendtskab med videnskabsfilosoffen Karl Popper. Det der umiddelbart gjorde et stort indtryk på mig, var hans klargørelse af et problem, der ofte (dette er selvfølgelig en efterrationalisering) har forvirret mig.
Hvordan kan vi være så sikre? - Er alle disse videnskabelige teorier egentlig beviste?.
At teorier er blevet fremlagte som beviste og sande har ofte naget min underbevidsthed, der vel har studset over tidligere sande teoriers undergang.
Popper løste (på en måde) induktionsproblemet, - at videnskab bygger på induktion (slutning fra enkelttilfælde til en almen lov) og at induktion intet kan bevise. Han viste den logiske forskel på verifikation (bevis) og falsifikation (modbevis), og gjorde falsificerbarhed til kriteriet på videnskabelighed.
(Det almindelige eksempel er det med svanerne. Man kan induktivt slutte, at alle svaner er hvide, udfra iagttagelser af en mængde svaner i Europa. Men fundet af en enkelt sort svane i Australien viser os, at vores slutning var falsk.)
(Magee 73, s 21)
Reelt er vi dog ikke kommet nærmere til 'sandheden' eller 'det endegyldige bevis'. For er det falsificerende udsagn gyldigt? Er det fortolket rigtigt? - Udsagnet og teorien skal omhandle 'det samme', spørgsmål og grænsedragninger skal være relevante osv. Falsifikationer er også underlagt fordomme og paradigmer.
Som Kuhn viste, har normalvidenskaben også metoder til at 'klare' det, der ligner falsifikationer. Vurderingen af, hvorvidt denne tilpasning af den bestående teori er berettiget eller ej, spiller en stor rolle i udviklingen af videnskabelige teorier.
(Kragh 81, s147)
Det afgørende i Poppers omformulering af induktionsproblemet ligger i, at hans 'løsning' giver en anden opfattelse af erkendelsesprocessen. Det vi kalder erkendelse og sandhed, også i form af videnskabelige teorier, må altid være noget foreløbigt!
"Den erkendelsesteori, jeg fremsætter, er altså stort set en darwinistisk erkendelsesteori. Fra amøben til Einstein er erkendelsens vækst altid den samme: Vi prøver på at løse vore problemer og på at opnå, gennem en eliminationsproces, noget der nærmer sig tilstrækkelighed i vore foreløbige løsninger." (Oversat fra Popper 73, s 261)
(Dette er for øvrigt kun ét blandt mange eksempler på brugen af biologiske og evolutionære analogier i filosofisk tænkning.)
1.5 Videnskabelig metode:
Poppers metodelære er forfriskende og sympatisk for mig, specielt fordi den viser nødvendigheden af åbenhed i videnskaben, og det kan være tiltrængt.
Den kendte fysiker David Bohm siger således:
"Det er ikke riktig som noen sier at vitenskapen starter med fakta. I virkeligheten tar vitenskapen utgangspunkt i en teori og stiller spørgsmål ut fra teorien. Bare det som besvarer disse spørgsmålene blir godtatt. Alle fakta som strider mot teorien blir ekskludert.
... Hvordan folk ser på bevis avhenger av hva de tror er mulig. Det de mener er umulig, vil de ikke ta alvorlig."
(Dammann 87, s 122)
Popper siger, at vi skal koncentrere os om problemer, - ja videnskab er for ham problemløsning. Problemer kan være alle slags 'skuffede' forventninger, lige fra de forventninger et stort teoretisk system formulerer om verden til de 'forventninger', der ligger i strukturen af vores sanseapparat.
Vi bør lave en foreløbig løsning til problemet, også selv om vi ved at den sandsynligvis er skrupforkert, denne løsning udsættes for (berettiget) kritik, og derigennem kan problemet undersøges udfra et lidt ændret grundlag. Gennem denne proces af foreløbige løsninger og kritiske tilbagevisninger forøges vores indsigt i problemet.
(Kragh 81, s 149f)
Darwin selv gav udtryk for en lignende opfattelse i 'Descent of Man':
"Forkerte data er højst ødelæggende for videnskabens fremskridt, for de består ofte længe; men forkerte opfattelser, med nogen støtte af beviser, gør sjældent skade, for alle finder fornøjelse i at vise de er forkerte."
(Her oversat fra Gould 83b, s 385)
Bekræftelser giver en nødvendig tiltro til den bestående teori, men de bidrager ikke til ny og forøget indsigt.
Kuhn hævder at falsifikation af den bestående teori lige så godt kan kaldes verifikation af den nye teori.
Jeg er ikke enig. - Hvis vi godtager, at videnskab er problemløsning, bliver den nye teori først til gennem denne problemløsning; en verifikation vil således altid gælde det bestående, det nye opstår først gennem falsifikationen af det gamle. Kuhn overser, at som proces vil en falsifikation være nyskabende, mens en verifikation vil være bevarende.
(Bl.a Kragh 81, s 168)
Gode videnskabelige teorier karakteriseres altså af deres falsificerbarhed eller prøvbarhed; - at de 'forbyder' en masse og let kan tænkes at komme i konflikt med konkrete observationer og eksperimenter.

(Fra Bonde 87, s 359)
Men Popper hævder vel at mærke ikke, at ikke-videnskabelige teorier efter disse kriterier er meningsløse, men anerkender i høj grad sådanne udsagns meningsfuldhed og værdi.
(Bl.a Kragh 81, s 147)
(Gould filosoferer i The Pandas Thumb over undtagelser i naturen, og når frem til at: 'besynderlige og sære ting er prøver på det generelle, ikke blot ejendommeligheder'.
Det engelske mundheld 'the exception proves the rule' (undtagelsen bekræfter reglen) udtrykker en almen sandhed, der er gået tabt i den sædvanlige forståelse. Det engelske proves stammer fra det latinske probare, der kan betyde både prøve og bevise. Mundheldet burde måske med større ret hedde: 'undtagelsen prøver reglen' på dansk.)
(Gould 83a, s 62)
En vigtig bemærkning til teorier og falsifikation er , at alle empiriske udsagn, også falske, har et sandhedsindhold (det er Poppers betegnelse for klassen af sande udsagn der følger af en teori). Ethvert falsk udsagn har uendelig (sic!) mange sande følger, og derfor kan en falsk teori føre os til en række sande konklusioner, og være overordentlig betydningsfuld og nyttig.
(Magee 73, s 38)
1.6 Bateson, natur og ånd:
Gregory Bateson har været af stor betydning for mig. Hver gang jeg læser hans bøger, synes jeg at han åbner mine øjne for ny erkendelse; desværre falder de alt for hurtigt i igen. Jeg synes, at han på mange måder harmonerer med Popper, og dog er han så anderledes i sit sigte og sine emner, at en sammenligning vel ikke er mulig.
Jeg tror Bateson dybest set var naturalist i samme forstand som jeg er det (se ovenfor), og følte at der i naturen og dens menneske ligger et grundlag for en etik, et udgangspunkt for en stillingtagen til hvordan ændringer burde foregå.
(Bl.a Ølgaard 86, s 138f)
Bateson lægger vægt på, at videnskabelige fremskridt ikke kan ske udelukkende gennem induktive studier af 'rå' data. Han understreger at 'data' ikke er begivenheder eller objekter, men altid registreringer, beskrivelser af eller erindringer om begivenheder og objekter. Der er altid en transformation eller omkodning af den rå begivenhed mellem videnskabsmanden og hans objekt.
Yderligere er der, altid og uundgåeligt, en udvælgelse af data, idet hele universet, før og nu, ikke kan observeres af nogen observatør. Iagttagelse er en selektiv proces!
En ensidig forkærlighed for induktion fører til hypoteser uden forbindelse til den fundamentale viden i videnskab og filosofi, og beskæftigelse med test af forudsigelser udfra disse hypoteser er udbytteløst mener Bateson, da det endelige mål i videnskab er forøgelse af den fundamentale viden.
Videnskabelig forskning må starte fra to udgangspunkter, observationerne kan ikke benægtes, og den fundamentale viden må anvendes.
En anden årsag til den manglende forbindelse mellem hypoteser, specielt indenfor biologi og adfærdsvidenskaberne, og fundamental viden, ligger i at en størsteparten af den grundliggende struktur i de forrige århundreders videnskab er uegnet eller irrelevant for problemerne og fænomenerne i disse videnskaber. Mentale processer, ideer, kommunikation, organisation og differentiation er emner af form og mønster snarere end af substans og energi, og i indledningen til Steps to an Ecology of Mind skriver Bateson afsluttende:
"Denne bog prøver at bygge en bro mellem livets og adfærdens kendsgerninger, og hvad vi i dag ved om mønster og orden." (Fra Bateson 85, s xv-xxvi)
Det vigtigste element i Batesons tænkning er kommunikationsteori, systemteori, kybernetik eller hvad man nu vælger at kalde det. Den videnskabstype der er vokset frem siden anden verdenskrig, og som omhandler organiserede helheder, og deres egenskaber som forandring og bevarelse, regulering og styring. Det der i mine øjne skiller Bateson ud fra andre systemteoretikere (ud over at han hører til blandt de klareste tænkere og formidlere jeg kender) er hans stadige fokuseren på det levende.
Det aspekt ved den levende natur, som Bateson koncentrerer sin interesse om, kalder han mind. Mind kan vel bedst oversættes ved ånd eller det lidt klodsede 'det mentale'.
Det er vigtigt her at lægge mærke til at mind eller det mentale er noget fælles for hele den levende natur, et mønster der forbinder. Dette betyder for Bateson også, at andre systemer end enkeltmennesker, fx dyr eller dyresamfund i én eller anden forstand kan tænke, erkende og få viden om verden og tingene.
Hvordan vi tænker, epistemologi eller erkendelsesteori, definerer Bateson således:
"En gren af naturvidenskaben forbundet med en gren af filosofien. Som naturvidenskab omfatter epistemologien studiet af, hvorledes særlige organismer eller systemer af organismer ved, tænker og beslutter. Som filosofi omfatter epistemologien studiet af de nødvendige grænser for og andre karakteristika ved de processer, der fører til at vide, tænke og beslutte."
Når epistemologi her defineres som problemet om, hvordan vi véd noget, så inkluderer Bateson i dette 'vi' også USA's senat, søstjerner, fyrreskove og æg. Og i det vi véd, indbefatter han alt, hvad levende væsener i én eller anden forstand véd noget om, fx hvordan de skal finde en mage, undslippe en fare, hvordan de skal vokse og stadig opretholde den samme form og ikke mindst, hvordan de skal udvikle sig.
(Ølgaard 86, s 9ff)
Jesper Hoffmeyer skriver i forordet til Batesons bog
Ånd og Natur:
"... nerven i hans sammenfatning er 'mønsteret som forbinder'. "Hvad er det for et mønster", spørger han et sted, "som forbinder en krabbe med en hummer og orkideen med kodriveren, og dem alle fire med mig? Og mig med dig? Og os alle seks med amøben i én retning og med den 'backward' skizofrene i en anden retning?"
Når vi ikke kan svare på det, er det måske fordi vi har misforstået, hvad ånd (mind) er for noget?
Når Darwins teori i sin tid vakte så megen rædsel, så var det i høj grad fordi den fjernede ånden fra naturen - og som vi kan se i dag fra mennesket. Der var ikke en hjerne bag naturens vidunder, men blind tilfældighed. Og det har mennesket lige siden haft hårdt ved at acceptere. Men hvad nu, hvis det åndelige hverken er noget spirituelt i sædvanlig plat forstand, eller noget, der nødvendigvis er knyttet til den enkelte menneskelige hjerne? Hvis det åndelige er et mønster, der forbinder? Hvis vi kunne finde det åndelige et andet sted, end vi tror, så ville det måske vise os enheden - vores enhed med en tusindfryd, eller med biosfæren for den sags skyld.
Bateson giver os kort sagt et andet begreb om det åndelige, om hvad det vil sige at være en 'ånd'. Og det fine er, at i og med, at det jordiske derved bliver åndeligt, så bliver det åndelige egentlig noget meget jordnært, eller naturligt. ...
"I det hele taget var det ikke de groveste, de simpleste, de mest dyriske og primitive aspekter af den menneskelige art, som blev afspejlet i naturens fænomener. Det var snarere de mere komplekse, de æstetiske, de intrikate og de elegante aspekter ved folk som afspejlede naturen. Det var ikke min begærlighed, min beregnende side, mine såkaldt 'dyriske' såkaldte 'instinkter' osv., som jeg lagde mærke til på den anden side af det spejl, derovre i 'naturen'. Snarere så jeg der rødderne af den menneskelige symmetri, skønhed og grimhed, æstetik, den menneskelige skabnings egentlige væren i live og dens smule visdom. Menneskets visdom, dets kropslige ynde og endog dets vane med at lave smukke genstande er lige så 'dyriske' som dets grusomhed. Når alt kommer til alt, så betyder selve ordet 'animal' (dyr) 'udstyret med ånd eller sjæl' (animus)." "
(Bateson 84, s 11f)
1.7 Videnskab, historie og myte:
Evolutionsteori har en meget speciel stilling i det videnskabelige system.
"Er nemlig evolutionsteorien på den ene side den ramme, uden for hvilken vi næppe har nogen chance for at forstå, hvorfra vi kommer, og hvori vi består, er det på den anden side også vigtigt at bestemme de grænser nøjere uden for hvilke den ikke længere fungerer som en videnskabelig teori, men som en myte."
(Jacob 85, s 15).
Evolutionsteorien omhandler fortiden, historien, og derfor er den ikke blot en naturvidenskabelig teori som alle de andre. Historien har jo den ubehagelige egenskab at den er sket.
Men evolutionen er også vores vigtigste myte, vores tilblivelseshistorie, og hvad dermed følger af implikationer for vores opfattelse af os selv.
Og derfor er teorierne omkring den blevet brugt og misbrugt i diskussionerne om, hvordan vi må behandle hinanden og resten af verden.
Om det videnskabelige siger Popper:
"Det, vi kalder udviklingshypotesen, er en forklaring af en mængde biologiske og palæontologiske iagttagelser - fx af visse ligheder imellem forskellige arter og slægter - ud fra den antagelse, at disse ligheder skyldes fælles afstamning. Denne hypotese er ikke en universel lov, selv om der ganske vist indgår visse universelle naturlove vedrørende fx arvelighed og mutation i den. Den har i stedet form af et individuelt, singulært historisk udsagn. (Den har samme form som 'Charles Darwin og Francis Galton havde samme bedstefar')."
Dette er dog ikke ensbetydende med at udviklingshypotesen ikke er videnskabelig, igen ifølge Popper, da singulære historiske udsagn godt kan være falsificerbare, en vævsprøve fra Darwin og Galton kunne fx udelukke, at de havde samme bedstefar.
Poppers indvending imod videnskabeligheden går i stedet på at selve evolutionsmekanismen er så vagt og tautologisk defineret, at der ikke er mulighed for at give forudsigelser af udviklingen og, at den derfor heller ikke kan falsificeres.
Tilpasning og selektion bliver anvendt tautologisk, - hvis en art er blevet elimineret, må den have været dårlig tilpasset og omvendt.
(Nielsen i Bonde 87, s 350)
Popper siger dog også, at selvom en teori viser sig at være ikke-videnskabelig, behøver den ikke at være betydningsløs eller meningsløs. Næsten alle videnskabelige teorier stammer historisk set fra myter, og myter kan således udvikles og gøres prøvbare.
(Magee 73, s 42).
Men man kan ikke udlede historiske love videnskabeligt ifølge Popper. Vi kan ikke finde en udviklingslov, der kan forklare hvorfor mennesket er som det er og forudsige hvad der sker fremover; naturen er ikke 'simpel', dens kompleksitet er ikke blot udtryk for nogle få grundlæggende love.
Evolutionsteoriens love og regler kan afstikke grænser for hvad vi mener der kan ske i en given situation, og derudover kan vi kun beskrive det historiske forløb.
Fransisco Ayala skriver herom:
"Der er to slags spørgsmål i studiet af biologisk evolution, den ene drejer sig om historie; studiet af fylogeni, udredningen og beskrivelsen af det faktiske evolutionære forløb på jorden, der har ført til den nuværende tilstand i den biologiske verden. ... Den anden slags spørgsmål drejer sig om forklaringen af den mekanisme eller proces, der bevirker evolutionær forandring. Disse spørgsmål omhandler årsager, snarere end historiske relationer."
(Oversat fra Dobzansky 77, s 486)
Problemet er at opdele spørgsmålene, oftest vil der i ethvert spørgsmål om evolution indgå både historiske og årsagsmæssige elementer.
Som Jacob skriver i citatet ovenfor, er det vigtigt at blive klar over hvad vi kan forklare videnskabeligt og hvad vi 'kun' kan beskrive, - eller forklare ved hjælp af myter.
På grund af evolutionens historiske og singulære natur står den i et vist modsætningsforhold til det traditionelle objektivitetsbegreb i videnskaben.
Ifølge den gængse videnskabsopfattelse er det hovedopgaven for respektabel videnskab at studere de egenskaber der er fælles for ting, idet disse formodes at give en dybere erkendelse af naturen end studiet af individuelle 'overfladefænomener'. Som følge heraf opfattes enkelteksemplarer kun som repræsentanter for en klasse, og abstraktion og generalisering er afgørende og nødvendige elementer i den videnskabelige proces.
(Kragh 81, s 215f)
Begrebet ensartethed - og dermed gentagelse - er derfor centralt i videnskab. Når vi siger, at nogle observationer er (fx induktivt bekræftende) gentagelser, er gentagelserne ikke eksakt identiske, men ligner blot hinanden i visse henseender, og det er os der definerer hvad der er lighed.
Den videnskabelige objektivitet opfordrer til at vi betragter lighederne og ignorerer forskellene, - vi vælger at se observationen som en gentagelse af en foregående observation.
Denne forkærlighed for det fælles udmønter sig ofte i, at abstraktionen og generaliseringen i sig selv forhindrer en større erkendelse af naturen som den er. Der udvikles en tro på, at komplekse fænomener kan erkendes udfra studier af simplere delfænomener. Om denne reduktionisme skriver Jesper Hoffmeyer:
"Det særlige ved naturvidenskaben er ikke, at den reducerer kompleksiteten, det særlige er den arrogance, den har udvist over for den ikke-viden, som reduktionen har forårsaget. Reduktionisme bør derfor ikke opfattes som selve den metode at gå reduktivt til værks i studiet af naturen. Begrebet bør i stedet bruges om den holdning at anse erkendelsestabet ved de fysiske reduktioner for ligegyldigt."
(Hoffmeyer 84, s 185)
Om begrebet ikke-viden vil jeg her blot sige, at det benyttes om de aspekter der bortabstraheres i reducerende forskningsmetoder, og om hvilke vi derfor véd, at vi ikke ved noget.
Stephen Jay Gould skriver om evolutionsteoriens vanskeligheder med det videnskabelige objektivitetsideal, at viden og forudsigelse ikke er det samme, og endvidere:
"Videnskaben om komplekse historiske emner er et anderledes, ikke et mindre, foretagende. Den prøver at forklare fortiden, ikke forudsige fremtiden. Den søger efter principper og regler under egenarten ved hver art og sammenhæng, mens den påskønner den irreducible egenart og beskriver al dens pragt. Videnskabens forestillinger må bøje sig (og udvides) for at passe til livet. Det løseliges kunst, Peter Medawars definition på videnskab, må ikke blive kortsigtet, for livet er langt."
(Oversat fra Gould 83b, s 65)
Kapitel 2 Indhold
INFORMATION OG KOMMUNIKATION
"Levende væsener kan kun overleve, vokse og mangfoldiggøre sig i kraft af en uafbrudt strøm af materie, energi og information."
(Jacob 85, s 109)
Stof- og energiaspektet ved levende væsener har længe været udgangspunkt for forskning og tænkning. Disse aspekter kan forklares og beskrives udfra den klassiske fysik.
Descartes mente, at dyrene var at ligne med maskiner (han sammenlignede dyrenes skrig under kniven med maskinstøj!), en forestilling der har været medvirkende til fysiologiens (lægevidenskabens) enorme udvikling og succes. Midt i dette århundrede fremkom nye og mere raffinerede maskiner, datamaskiner, der kunne behandle information. Den videnskab der opstod under udviklingen af disse kaldes kommunikationsteori eller informationsteori, og nu blev man selvfølgelig (!) opmærksom på også dette aspekt ved de levende 'maskiner'.
Gregory Bateson var med fra begyndelsen af kommunikationsteoriens udvikling, og specielt han brugte den i høj grad på nye områder som liv, udvikling og overlevelse.
2.1 Kommunikation og sprog:
Det fascinerende og det vanskelige ved kommunikationsteori er, at den omhandler et helt andet aspekt ved verden end de materielle, stoflige og energimæssige aspekter. Den anvender derfor også et helt andet begrebssprog end det klassiske newtonske, der 'blot' er en præcisering af vores dagligsprog, fundamentet i vores indbyrdes kommunikation om det abstrakte.
Vi taler til daglig som om en ting kunne 'have' en eller anden egenskab; stenen er hård, dyret er lille, osv.
"Vil man tænke klart er det tilrådeligt at forvente, at alle egenskaber, attributter, adjektiver og så videre refererer til mindst to sæt af samspil." siger Bateson. Stenens modstand mod indtrængen - og dens indre sammenhæng. Dyrets størrelse i forhold til den talende - og i forhold til andre dyr af dets egen art, køn og alder. "Det ville være en mere præcis talemåde at sige, at 'tingene' er fremstillet, gjort 'virkelige' i kraft af deres indre forhold og deres adfærd i forhold til andre ting og den talende."
(Bateson 84, s 56f)
Uanset hvad 'tingene' er i deres tingslige verden, kan de kun omtales og få mening udfra information om deres indre og ydre forhold og samspil; tings egenskaber er altid egenskaber i en bestemt sammenhæng. Og denne information kategoriseres udfra vores intersubjektive (fællesmenneskelige) antagelser og normer, hvoraf en del ligger i sproget.
Se også Peter Zinkernagels bog: Omverdensproblemet (også i Nørretranders 85, s 237ff), for en analyse af dagligsprogets betydning i vores forståelse af omverdenen og i et bevis for, at det er meningsløst at fremføre den ontologiske påstand at verden ikke er til udenfor vore tanker. - Og Bohm 86, s 43ff, hvor han undersøger hvorledes sproget bidrager til tankens fragmentering, til at opdelinger spiller så stor en rolle i vores kommunikation.
Bateson siger at kommunikationsteorien omhandler det emiske aspekt af ting, hændelser og adfærd; dvs. den information, der udsendes (emitto = udsende) eller 'bæres' af tingene, hændelserne og adfærden. Hvis en tyr står foran dig og skraber med det ene forben i jorden, er hverken tyren eller du interesseret i den bevægelse af jord og græs der sker, her er det kun det emiske aspekt, informationen der er af betydning.
Information definerer Bateson som en forskel der gør en forskel (altså opfattes). Heri ligger kimen til hvorfor de newtonske begreber ikke kan anvendes på kommunikation, - en forskel er dimensionsløs og kan ikke lokaliseres i rummet.
(Ølgaard 86, s 44ff).
Årsag og virkning i det materielle univers kan beskrives som én del der påvirker en anden del, via en eller anden form for 'kraft'. Hvad kommunikation angår kræves der derimod et forhold, enten mellem to dele eller mellem en og samme del på to tidspunkter, (- altså en forskel eller en forandring) for at aktivere en tredje del, der kan kaldes modtageren. Det vigtige led her er naturligvis modtageren, uden hvilken information kun findes i en potentiel eller uvirkelig form, som en uendelig mængde af mulige forskelle.
Rør ved et glas med koldt vand og et med varmt. Forskellen mellem kold og varm findes hverken i det ene eller det andet glas, eller i rummet imellem dem. Men dog opfatter du forskellen, og den bliver til information i dig.
Desuden kan du som menneske præcisere denne forskel, da den kan omformes til en kvantitativ forskel, dvs. en forskel der kan beskrives i rum og tid (i dette tilfælde forskellen på længden af en kviksølvsøjle i et glasrør). Kvalitative forskelle er forskelle vi endnu (?) ikke kan omforme på denne måde.
Videnskabsmænd søger ofte efter måder at omforme kvalitative forskelle på, således at de kan beskrives i rum og tid.
Dette skyldes måske, at rum/tid-begreber er udtryk for en (intersubjektiv) måde at opfatte genstande og hændelser i vores omverden på, der er iboende i vores sanseorganer i så stor grad, at rum og tid føles lige så 'virkelige' som genstandene og hændelserne.
Jeg mener ikke at genstande og hændelser er mere 'virkelige'. Min pointe er blot, at når vi sanser noget, er det ikke et 'virkelighedskriterium', at det kan beskrives i rum og tid, - uanset nytteværdien af en sådan beskrivelse. Jeg argumenterer egentlig for at kvalitative forskelle er lige så vigtige og vægtige som kvantitative, uanset om de er vanskeligere at have med at gøre.
Kvalitet er et uomgåeligt begreb i en bedre forståelse af verden. Grundliggende forskelle i ånd og videnskab er kvalitative; at 'vælge' imellem dem er at tillægge dem værdier. Værdier er udtryk for en eller anden form for anvendelighed, en vis overensstemmelse med omverdenen.
Et valg mellem to grundliggende kvaliteter som fx den euklidiske og den riemannske geometri (der er mange andre geometrier), er principielt som et valg mellem at måle i meter eller favne. Et geometrisk aksiom (eller andre a priori syntetiske 'udsagn') kan ikke falsificeres, men blot gå ud af brug, hvis det ikke længere er praktisk. Begge geometrier er 'sande' i den forstand, at de er uden indre logiske modsætninger, og en modsigelse af den ene med den anden er ikke mulig (i relativitetsteorien er den riemannske for øvrigt mest anvendelig).
Ovenstående er bl.a inspireret af Robert M. Pirsig; jeg synes at hans bog er en af de sjældne, der føles som en personlig oplevelse, når man læser den.
(Bl.a Pirsig 85, s 236ff)
2.3 Lidt om sansning:
Det fremgår af det foregående, at information er knyttet sammen med sansning, eller perception.
Fritsvømmende mikroorganismer vil kunne percipere informationen i en næringsgradient, og bevæge sig selektivt langs denne gradient, mens fx en flamme ikke har denne evne, men blot ernærer sig og vokser proportionalt med den tilgængelige energi, flammen vil ikke bevæge sig selektivt.
Hvis vi ser en hjort der står fuldstændig stille, er det ikke umiddelbart klart hvilke forskelle der egentlig giver os information om dyrets form. Står vi selv stille vil hjorten (og de fleste andre dyr) ikke 'se' os, den opfatter os ikke før vi bevæger os og derved skaber forskelle i hjortens øje. Grunden til at vi kan se hjorten selv på et sort/hvidt billede er, at vores øje hele tiden laver små bevægelser kaldet mikronystagmus; det vibrerer nogle få buesekunder og 'skaber' derved konturerne i vores omverden via de forskelle der opstår i de lysfølsomme tappe og stave.
'Gradvise' eller 'langsomme' forandringer er vanskelige at opfatte, fordi de maskeres af det fænomen der hedder tilpasning eller tilvænning (fordi de normalt er uden interesse?).
Bateson giver et eksempel, jeg ikke kan lade være med at citere:
"En halvvidenskabelig fabel fortæller, at hvis man kan få en frø til at sidde stille i en kasserolle med koldt vand, og man så opvarmer vandet langsomt og jævnt, så der ikke forekommer et øjeblik, der markeres som det øjeblik, hvor frøen bør springe, så springer den aldrig. Den bliver kogt. Er mennesket i færd med at ændre sit eget miljø med langsomt voksende forurening og sit åndelige system med langsomme forringelser af religion og uddannelse i sådan en kasserolle?"
(Bateson 84, s 90ff)
Jeg vil ikke gå videre med perception her, men blot endnu engang understrege, at alle levende væseners måde at opfatte omverdenen på (deres epistemologi) er rodfæstet i deres perceptionsudstyr og -processer, i redskaberne for skabelse og behandling af forskelle.
Francois Jacob skriver om dette:
"For hver art afhænger den ydre verden, sådan som den perciperes, dels af sanseorganerne, dels af den måde, hvorpå hjernen integrerer sensoriske og motoriske begivenheder. Selv når forskellige arter perciperer en og samme skala af stimuli kan deres hjerne være organiseret således, at forskellige særegenheder udvælges. Miljøet kan, alt efter hvordan informationen behandles, afvige lige så radikalt, som hvis de opfattede stimuli hidrørte fra forskellige verdener. Vi er selv så snævert indespærrede i den afbildning af verden, som vort sanse- og nerveudstyr påtvinger os, at vi har svært ved at tænke os, at det er muligt at se denne verden på nogen anden måde. Vi kan vanskeligt forestille os en flues, en regnorms eller en måges verden."
(Jacob 85, s 111)
Hvis man vælger at opfatte og forstå verden (også) udfra det emiske aspekt, ser man, at vi alle er viklet ind i et netværk af information og kommunikation. Kommunikation er indeholdt i det vi forstår ved liv, fra det højeste til det laveste. Bateson går så langt som til at sige, at:
"a priori kan man hævde, at al perception og al respons, al adfærd, al indlæring og al genetik, al neurofysiologi og endokrinologi (hormoners funktion og virkning), al organisation og al evolution - ét eneste emneområde - må betragtes som kommunikatorisk af natur og derfor genstand for de store generaliseringer eller 'love', som gælder for kommunikatoriske fænomener."
(Her fra Ølgaard 86, s47)
Processer, ligegyldigt af hvad art, kræver energi, og informationsprocesser er ingen undtagelse.
Men informationsprocesser adskiller sig alligevel fra 'almindelige' fysiske processer.
Fysiske processer er karakteriseret ved at energien (næsten) altid er intern eller 'bagfrakommende'. Dvs. at den fysiske virkning et fysisk objekt forårsager, altid kan forudsiges udfra viden om objektets og 'det påvirkede objekts' stoflige og energetiske karakteristika.
Forskellen til informationsprocesser ligger faktisk i afgrænsningen af 'det påvirkede objekt', her er 'virkningen' nemlig også afhængig af ekstern eller sideordnet energi, dvs. energetiske karakteristika, der ikke ville blive inddraget i en fysisk beskrivelse (jeg kan ikke definere det mere præcist, da der ikke er tale om fænomener der ligger udenfor fysiske beskrivelser, men om en anden synsvinkel eller netop et udvidet fokus).
Hvis vi knipser til en sten, kan vi finde en ret præcis sammenhæng mellem stenens størrelse, kraften i knipset og den observerede effekt.
Knipser vi derimod til et blad på en følsom mimose, afhænger effekten også af den indre potentielle energi i mimosens motoriske celler og endda af den tilstand af tilvænning mimosens sanseapparat befinder sig i. Hvor den indre potentielle energi kan sammenlignes med vandtrykket når en vandhane åbnes, er de energetiske forhold i sanseapparatet langt sværere at beskrive (billedet af informationsbehandlende relæer i en dataprocessor, med signalstrøm og styrestrøm, intern og ekstern, kan være nyttigt for at forstå de basale principper; men næppe af nogen værdi for en forståelse af hvad der egentlig sker).
Når det kommer til de processer der afgør fx, hvordan en hund reagerer når den møder en anden hund eller, hvordan en mand reagerer på et slag på kinden med en handske, er vores viden om de involverede processer meget begrænset.
(Inspireret af Ølgaard 86, s 146f)
2.5 Tilfældighed og valg:
Vi støder her på et aspekt af spørgsmålet om eksistensen af 'frie valg', forstået på den måde at en proces, der forløber i et meget komplekst system, ikke kan være determineret (forudbestemt) efter min opfattelse, men må have et indhold af tilfældighed.
Det er vigtigt her at skelne mellem tilfældighed og kaos, man kan ikke kaste terninger med glaskugler. Tilfældigheder er tilstede og synlige, netop fordi verden for det meste er determineret. Terningekastet er ikke kaotisk, men består i distinkte udfald, grundet i form, struktur og massetiltrækning, der er fordelt på en tilfældig måde.
(Karl Pribram, her fra Dammann 87, s 259ff)
Tilfældigheden udgør grundlaget for et egentligt valg.
Et valg forudsætter først og fremmest at der findes valgmuligheder, på samme måde som et terningekast forudsætter 'valgmuligheder' (udfald). Der findes 'tilfældige valg' der, præcis som (og måske ved hjælp af) et terningekast, 'afgøres' af det underliggende kaos, sådanne 'valg' forekommer når forskellen mellem valgmulighederne ikke erkendes.
Et valg kan kun kaldes et frit valg, hvis erkendelsen af en forskel i værdi, nyttighed, rigtighed eller lignende, kan resultere i valget af det, der er af mindst værdi, nyttighed eller rigtighed. På den anden side er disse og lignende begrebers indhold betinget af eksistensen af (eller forestillingen om) et frit valg; ligeledes har begreber som ansvar og etik ingen mening uden det frie valg og den bagvedliggende indeterminisme.
Årsagen til at jeg kommer ind på indeterminisme her, er at jeg vil antyde hvorfor en fastholden af det traditionelle fysiske verdensbillede og de antagelser der ligger heri, vil vanskeliggøre forståelsen af bl.a det emiske aspekt, og dermed det levende som helhed, på tilfredsstillende måde.
Det deterministiske verdensbillede kan vanskeligt forenes med kvantemekanikkens beskrivelse af verdens meget små dele, og den erkendelse der ligger heri.
Det har gennem længere tid stået uimodsagt, at hændelser af kvantemekanisk størrelsesorden ikke er præcist 'bestemte', men indeholder et vist mål af tilfældighed. Werner Heisenbergs ubestemthedsrelationer formulerer nogle aspekter heraf. Kvantemekanikken kommer derfor også i modstrid med den klassiske opfattelse af objektivitet i videnskaben, som jeg har været inde på i forrige kapitel.
Grunden til at jeg omtaler kvantemekanikken her er ikke, at jeg tror, at de forestillinger og antagelser, der ligger i den nuværende kvantemekaniske teori, nødvendigvis er 'rigtigere' end de newtonske, men netop for at belyse, at sådanne antagelser findes og er af betydning for vores verdensbillede.
Niels Bohrs forslag til et nyt objektivitetsbegreb passer faktisk på mange måder bedre til biologisk videnskab, end det klassiske objektivitetsbegreb som jeg omtalte i afsnit 1.8.
Bohrs objektivitetsbegreb er tilpasset den kvantemekaniske situation, at eksperimenternes udfald hænger nøje sammen med forsøgsopstillingen, og at samme forsøgsopstilling i almindelighed giver forskellige resultater, pga. den tilfældighed der er indeholdt i situationen. Det afgørende grundlag for denne objektivitet er de erfaringer, der entydigt kan meddeles til andre, uafhængigt af det iagttagende subjekt. I denne entydighed ligger at hele iagttagelsessituationen må beskrives og indgå i definitionen af det iagttagne fænomen. Fænomenet 'i sig selv' kan ikke defineres objektivt, da skillelinjen eller grænsen mellem fænomen og det instrument, der indgår i iagttagelsen, ikke er 'bestemt'; vi afgør hvor vi vil sige at grænsen er i den givne situation.
En konsekvens af denne uadskillelighed mellem fænomen og iagttagelse er, at den erkendelse om fænomenerne vi får fra de forskellige eksperimenter, ikke bliver en éntydig erkendelse af fænomenerne.
Bohr opnår alligevel en entydig beskrivelse ved hjælp af det han kalder komplementaritetsprincippet.
Fx kan (og må) en foton eller en elektron betragtes og beskrives på to hinanden udelukkende men hver især utilstrækkelige måder, to komplementære måder, en bølgemåde og en partikelmåde. En elektron kan fx udvise bølgeinterferens (også med 'sig selv'!) og registreres diskret som en partikel i det selvsamme forsøg (Bohrs dobbeltspalteforsøg).
(Bl.a Kragh 81, s 222ff)
Som ovenfor nævnt har Bohrs forslag til et nyt objektivitetsbegreb i fysikken konsekvenser for opfattelsen af videnskabelig erkendelse i helt andre videnskaber, som biologi og sociale videnskaber, der har krympet sig under det klassiske ideals monopolisering af videnskabelig erkendelse. Det er måske også givtigt at forsøge at tænke i komplementære forståelsesveje omkring fænomener som intuition og logik, analyse og oplevelse, reduktion og helhedsforståelse, mv.
(Bl.a Kragh 81, s 240)
Min hensigt med ovennævnte bemærkninger er også at minde om, at kvantemekaniske processer er grundlaget i levende organismer. De ændringer i forståelsen af verden kvantemekanisk tænkning forårsager kan ikke ignoreres. Vi har fået et nyt grundlag for at forstå vores verden, og tilfældighed eller ubestemthed er et af de mest fundamentale aspekter heri.
2.7 Den virkelige verden:
Jeg vil ikke udelade det mest revolutionerende og aktuelle i kvantemekanikken, nemlig løsningen af et paradoks som Albert Einstein formulerede sammen med fysikerne Podolsky og Rosen i 1935. Paradokset var, at kvantemekanikkens konsekvenser var i modstrid med den virkelighed, de tre fysikere (og næsten alle os andre) mente at kende.
Det såkaldte Aspect-eksperiment er i 1981-82 (og mange gange senere) forløbet som kvantemekanikken og Bohr forudsagde. Dette eksperiment skulle påvise, at produkter af kvantemekaniske hændelser, har en forbindelse med hinanden udenfor rum og tid. - Eller egentlig skulle forsøget afvise at der skete ting, der ikke kunne forklares indenfor den gældende virkelighedsopfattelse, og således falsificere dele af den kvantemekaniske teori.
Det følger som yderste konsekvens af eksperimentet og kvantemekanikken, at en fotons (lyspartikels) skæbne i én galakse hænger sammen med dens tvilling i en anden galakse.
Jeg er ikke i stand til at vurdere eksperimentet som sådan, og fortolkningen af kvantemekaniske eksperimenter er som ovenfor nævnt ikke ligetil; men det, at problemstillingen siden Einstein og Bohrs diskussioner har været diskuteret af fysikere (både de der forventede en bekræftelse og de der ventede det modsatte) og, at eksperimentet betragtes som det måske vigtigste i dette århundrede, siger noget om de voldsomme konsekvenser denne del af kvantemekanikken har for vores forståelse af verden.
(Bl.a Dammann 87, s 39ff)
Det faktiske forsøg udførtes ved, at calciumatomer blev 'anslået' af laserlys. Calciumatomerne udsender derpå to 'tvillinge'-fotoner, der registreres efter at de har passeret gennem et polariseret materiale (som i polaroidsolbriller), der bremser fotoner med modsat polarisering. De fotoner der udsendes har samme polarisering.
Einstein ville sige, at polariseringen er en egenskab ved partiklen, fra det øjeblik de to fotoner skilles, og at de to fotoner derfor eksisterer uafhængigt af hinanden. Kvantemekanikken siger, at en partikels polarisering først bliver 'en realitet' i selve målingen, men stadig må fotonernes polarisering være den samme. Bohr fortolkede kvantemekanikken således, at parret må udgøre en sammenhørende totalitet, selvom de er lysår fra hinanden.
Det synes umuligt at skelne mellem de to opfattelser eksperimentelt. Men John Bell viste at graden af sammenhæng (korrelation) mellem adskilte systemer ikke kan overskride en vis grænse, hvis det er som Einstein og fornuften siger. Hvis kvanteteorien har ret, kan denne grænse passeres. Intuitivt forstår jeg dette udfra, at selvstændige eksistenser kan forstyrres uafhængigt af hinanden, hvorved korrelationen kan ødelægges, mens en stadig samhørighed vil bevare korrelationen.
Hvis de polariserede plader er parallelle, vil begge fotoner enten blive registreret eller ej. Men hvis de står lidt skævt for hinanden, vil der kun være en vis sammenhæng mellem registreringen.
Det målte sammenfald i eksperimentet er så stort, at det ikke kan forklares udfra den opfattelse at partiklerne eksisterer hver for sig. Som Bohr sagde, må der være en samhørighed mellem fotonerne. Samhørigheden kan ikke forklares som en slags kommunikation i (det normale) rum og tid, da de to fotoner bevæger sig med lysets hastighed.
(Davies 87, s 128ff og Nørretranders 85, s 155ff)
2.8 Opdelinger og grænser:
Som jeg før har nævnt er gentagelser og ensartethed ikke noget 'naturgivent', men konsekvenser af de valg vi foretager, opdelingen af vores verden i enheder kan foretages på mange måder.
Set udfra energiudveksling og fysisk/stoflig sammenhæng, er de iøjnefaldende grænser cellernes ydre membraner og individernes overflade; celler og individer er energetiske og stoflige enheder.
Betragter man længere tidsrum end individernes livslængde støder man på intime forbindelser mellem individer, og enheder som familier, populationer eller avlslinier dukker op. Føres sådanne tanker videre, fremkommer mere hypotetiske enheder, der bygger på muligheden af, at individerne kan danne udgangspunkt for familier, arten er en sådan enhed. Og ikke mindst opstår tanken om evolution, at alt levende på denne måde er forbundet i tid; og dermed mere vilkårligt valgte enheder (bl.a proportionalt med den tidslige afstand) som slægter, ordener og lignende.
Hvis man i stedet for den tidslige udstrækning fokuserer på den rumlige, finder man yderligere forbindelser af energi og stof, og enheder som fødekæder, fødenet og økosystemer viser sig.
I evolutionsteorien har man diskuteret begrebet evolutionsenheden udfra den opfattelse, at visse evolutionsmekanismer antages at 'virke' på og forandre en bestemt enhed i naturen, og derigennem bevirke de forandringer, der findes afspejlet i geologiske fakta og biokemiske, anatomiske, fysiologiske og adfærds- ligheder.
Jeg vil senere gå nærmere ind på problemerne heri, og blot her pege på, at grænserne for en sådan enhed hidtil er blevet fastlagt udfra en energetisk/stoflig opfattelse af enheder i naturen og, at en inddragelse af emiske aspekter givetvis vil have indflydelse på vores opdelinger af den levende verden.
Forestil dig en mand der fælder et træ med en økse.
Din forestilling indeholder formentlig, som det ligger i sprogets opbygning, tre enheder, hvor den ene er aktiv og 'bestemmer' eller kontrollerer situationen, og den anden passiv. Den tredje er et redskab, en slags forlængelse eller udvidelse af den aktive del.
Bateson ser et cirkulært kredsløb; - forskelle i veddet, hvor øksen lander, bliver til forskelle i mandens nethinde, forskelle i hans centralnervesystem, i hans motoriske nerver, i hans muskler, i øksens bane, i veddet, hvor øksen lander, osv., - et kybernetisk system.
Du vil måske mene, at der stadig er tre klare enheder, uanset at man kan betragte et informationskredsløb igennem dem. Et andet eksempel viser at enhederne ikke altid kan opretholdes, hvis man skal forstå hvad der sker.
Du er nu en blind mand, der går langs et fortov ved hjælp af en stok (sådan siger vores sprog - en alternativ beskrivelse virker kluntet og kompliceret, netop fordi sproget indeholder mange af vores fælles antagelser eller fordomme om omverdenen). Hvor går grænserne for dit 'jeg'? Er det afgrænset af huden, hører håndtaget med, eller hele stokken? Hvis du tror det er simpelt at afgøre, bør du gøre forsøget. Finder du, at du ikke kan skille stokken ud fra dig selv, prøv da at løsne grebet om håndtaget og du vil pludselig føle at stokken er 'noget du rører ved', hvor du før 'rørte ved noget med stokken'. Begge opfattelser er indeholdt i situationen. Den enhed der giver den bedste forståelse er manden-og-stokken-og-fortovet, med det informationskredsløb enheden indeholder. Når manden spiser er enheden en anden.
De afgrænsninger vi skal foretage for at forstå virkeligheden, er langt fra så simple og konstante, som når vi forsøger at forstå den mere indskrænkede virkelighed, hvor det emiske aspekt er bortabstraheres.
Det 'system' man betragter må afgrænses således, at de relevante kredsløb ikke skæres over men indbefattes i systemet, hvis vi skal kunne forstå hvad der sker.
(Bateson 85, s 448-466)
2.9 Beskrivelse, forklaring og forståelse:
Jeg har brugt ordene forklaring og især forståelse meget; det er måske rimeligt at forsøge at belyse de begreber der ligger bag ordene.
Hvad er forskellen på at beskrive en hændelse eller et fænomen, og at forklare dem? Spørgsmålet er mere berettiget end det umiddelbart lyder.
Begge begreber bruges om kommunikation om eksempelvis et vist fænomen, men en forklaring formodes at give større forståelse end en 'ren' beskrivelse.
En beskrivelse er et forsøg på at omforme information om fænomenet på en meget analog, lidet forvrængende måde, således at flest muligt af de sansede, virkende forskelle om fænomenet overføres til modtageren. Beskrivelsens effektivitet begrænses af den matrice eller 'støbeform' i hvilken kommunikationen foregår. Et landskab kan fx beskrives fotografisk, det kan tegnes på et stykke fladt papir, i et plastikrelief, på en globus, og det kan beskrives sprogligt på et utal af måder, i digt eller prosa, mundtligt eller skriftligt. Hver matrice har sine fordele og begrænsninger i formidlingen af det sansede.
Forklaringen er på samme måde begrænset af den benyttede matrice, der ikke behøver at være den samme som for beskrivelsen. Den grundliggende matrice i al kommunikation kan siges at være beskriverens og modtagerens epistemologier, deres måde at percipere, sanse, opfatte og tænke om verden på, deres 'verdensbilleder'.
En forklaring tilfører ikke ny information udover den der findes i en 'fuldstændig' beskrivelse. Faktisk forsvinder en stor del af informationen i beskrivelsen ved en forklaring, ja - netop dette er en vigtig del af forklaringen, at udelade de for den ønskede forståelse irrelevante dele af beskrivelsen, og dermed fremhæve de relevante.
Udover dette karakteriserer Bateson forklaring som en kortlægning af en beskrivelse ved hjælp af en tautologi, en række formelt eller logisk gyldige sammenhænge mellem de udvalgte forudsætninger i beskrivelsen.
At en forklaring kan give indsigt og forståelse, må således også ligge i udformningen af tautologien.
Vores forklaringer indeholder sjældent strengt logiske tautologier, som i matematikken; snarere er gyldigheden af tautologiens led betinget af en fælles accept, af om leddene er 'selvindlysende' udfra 'sund fornuft' eller lignende antagelser der er mere eller mindre fælles.
Accepten af tautologierne bygger ofte på associationer til andre fænomener hvor lignende sammenhænge forekommer, på analogier til andre fænomener.
(Bateson 84, s 77ff)
Det betragtes da også som en støtte for rent videnskabelige hypoteser, hvis hypotesen er udtryk for en forklaringsform, der er analog til allerede accepterede teorier.
(Kragh 81, s 40f)
Jeg betragter associationer til analoge sammenhænge i mere velkendte fænomener som noget uhyre vigtigt for forståelsen af nye fænomener og problemer. Jeg ser faktisk ikke andre veje til at forstå noget nyt.
Er det nye fundamentalt forskelligt fra det kendte, vil de associationerne der dukker op næppe være anvendelige, - fænomenet vil være uforståeligt indtil gabet mellem det nye og det kendte kan indsnævres, og en forbindelse kan ses.
2.10 Noget om sansning, genkendelse og forståelse:
Det er muligvis nyttigt at prøve at udvide området for overvejelserne.
Det følgende indeholder viderereflektioner over bl.a Batesons tanker om perception, som jeg var inde på tidligere i kapitlet.
Jeg bruger her 'perception' om selve processerne i sanse-organerne, mens 'sansning' bruges i en bredere betydning i retning af erkendelse og forståelse.
Sansning kan siges at bestå af flere led af forskellige typer af sammenligning og genkendelse.
I perceptionen bliver de fysiske processer sammenlignet med organismens perceptionsapparat; visse former stemmer bedre overens med (genkendes lettere af) sanseorganerne end andre. Mennesker vil fx percipere synligt lys i langt større omfang end anden elektromagnetisk stråling.
Derefter sker der det, at påvirkningen af stråling eller lignende 'sammenlignes' med den umiddelbart forudgående påvirkning, det er derfor at perception sker i form af forskelle. Det kommer her til syne, at disse perceptionens forskelle altid er forskelle i tiden, forandringer.
Næste led er en sammenligning af noget der ligner relevans eller overensstemmelse, den information der opfattes eller genkendes (i en eller anden forstand) er 'relevant' for organismen. Her må genkendelsen bygge på en mere kompleks form for hukommelse end de rent mekaniske 'hukommelser' ovenfor. Der må være en kobling mellem hukommelsens mekanismer og formåen, og vanskelighederne i at skelne mellem konstans og langsomme kontinuerte forandringer. Forskelle, der ikke indeholder tid, bliver formentlig til, bliver sanset ved hjælp af en sådan hukommelse, - såsom farver, rumlige berøringsforskelle og lignende.
Forståelse kan ses som et lignende led, en sammenligning af ny information med information fra kendte sammenhænge, enten fra vores egen interne hukommelse eller fra en hukommelse udenfor, hvor forståelsen er den genkendelse eller sammensætning af genkendelser man oplever.
Forklaring må da være en, til dette formål, særlig nyttig form for ekstern hukommelse.
Genkendelsen må (som en gentagelse) være et slags 'valg', et valg mellem at 'se' ligheder eller forskelle, lige fra det meget håndfaste valg i perceptionen til de mere subtile valg i fænomener som erkendelse og association. Valget synes at være en afgørelse af om noget der ligner relevans, interesse, harmoni og overensstemmelse er til stede eller ej.
Det er min fornemmelse at dette valg udgør noget analogt for processerne i sansning.
Bateson nævner et fænomen, der er af interesse her, han kalder det moiré-fænomenet (fra fransk, et silkestof hvor vævning og en speciel presning giver et mønster der ligner årer i træ).
Fænomenet opstår når mønstre mødes eller kombineres. Et meget enkelt eksempel er de stød, der opstår når to lyde med forskellig frekvens mødes, fænomenet udnyttes fx når man stemmer musikinstrumenter.
Men hvad er mønstre egentlig?
Mønster og mængde (her i betydningen kvantitet) er af forskellig logisk type (se afsnit 4.2), et mønster kan indeholde en eller flere mængder, men én mængde kan ikke fastlægge eller indeholde et mønster. Eller - et mønster består i forhold eller relationer mellem to eller flere mængder, ikke i disse mængder selv. Mønstre kan være overordentlig komplekse, indeholde uhyre mange relationer af flere logiske typer.
(Bateson 84, s 75f og 41ff)
Man kan også sige at mønstre er sammensætninger af forskelle, i hvilke der findes redundans (overflødighed eller gentagelse). Det mønster kvantiteterne danner, består af redundansen, af regelmæssigheder, orden eller begrænsninger i kaos. Redundansen er det, de forskellige forskelle 'siger om' hinanden.
Hvis kvantiteterne er 'tilstrækkeligt ens' (hvis der findes redundans), vil relationerne imellem dem altså udgøre et mønster. Ligeledes vil mønstre der stemmer mere eller mindre overens danne et nyt mønster, hvis de mødes; dette mønster indeholder relationer mellem de oprindelige mønstre (og er derfor af en anden logisk type).
Interferensmønstre er sådanne mønstre, på et simpelt fysisk plan. Det er fx et interferensmønster, mellem to laserstråler, hvor den ene er reflekteret fra motivet, der ligger på en holografisk plade.
Afgørelsen af om noget der ligner relevans, interesse, harmoni og overensstemmelse er til stede eller ej mellem mønstre eller kvantiteter, synes at indgå på mange niveauer i sansningen. Jeg tror at opmærksomhed på moiré-fænomenet vil være af værdi her og andre steder, hvor mønstre af enhver art mødes.
Bateson nævner kort at fænomenet muligvis kan bidrage til forståelse af økologiske nicher, æstetiske oplevelser og bevidstheden, men går ikke videre.
En forklaring kan altså opfattes som - at bringe det nye mønster (beskrivelsen) på en sådan form (ved udvælgelse af visse dele og knytning af visse forbindelser frem for andre), at det vi kalder forståelse kan opstå, måske ved 'noget der minder om moiré' med den forståelse vi allerede har. Et nyt 'mønster' vil altså kun kunne forstås i det omfang sådanne varierende forklaringer kan bringe det på en form, der stemmer overens (danner redundans) med vores øjeblikkelige erkendelse - og næppe nogensinde 'fuldt ud'.
Den nye forståelse der opnås, kan på den anden side ses som en slags harmoni mellem ny information og den allerede opnåede erkendelse. Den erkendelse eller viden vi (eller andre levende væsener) besidder, er så lig med det omfang, vi er i overensstemmelse med de mønstre vi møder, - og derfor relativ efter situationen.
Indeholdt i ovenstående overvejelser over hvad forklaring og forståelse er, ligger måske netop grundlaget for det, jeg i det foregående har kaldt fordomme eller paradigmer i dagligdag og videnskab. I forklaringens natur ligger, at uden en overensstemmelse i visse grundliggende antagelser er muligheden for at forklare, begrunde eller formidle forståelse meget små.
I den mere formelle videnskabsteori bygger Erlangerskolen (eller den konstruktive videnskabsteori) på lignende principper for videnskabelig begrundelse.
Det formuleres på den måde, at videnskabelige påstande skal kunne forsvares i en rationel dialog; det rationelle henviser her til en fælles fornuft dvs. et fælles sprog, og muligheden for fælles accept af gyldigheden af en dialogs påstande.
Igennem denne dialog kan en videnskabelig påstand føres tilbage til simplere påstande, der kan afgøres af 'praksis', dvs. en førvidenskabelig adskillelses- og orienteringsform.
Erlangerskolens praksis tilhører den type udsagn der kaldes syntetiske a priori, dvs. et udsagn om virkeligheden, som siges at være gyldigt uafhængigt af vores erfaring. Jeg mener at denne praksis er noget i retning af den fællesmenneskelige epistemologi, grundet i vores fælles sanse- og erkendelsesapparat.
(Kragh 81, s 74ff)
Kapitel 3 Indhold
EVOLUTION
Evolutionstanken fandtes længe før Darwin, men publikationen af 'Arternes oprindelse' i 1859 står som et vendepunkt for argumentationen omkring evolution. Darwin havde haft over tyve år at overveje argumenterne siden den centrale ide om 'naturlig udvælgelse' slog rod i ham. Bogen indeholder da også et væld af eksempler og argumenter, der afprøver ideen om en evolution hovedsageligt baseret på naturlig udvælgelse. Dette overbeviste de fleste videnskabsmænd om, at den levende verden er blevet til gennem en evolution, men ironisk nok sjældent om den centrale mekanisme (oftest pga. modstand mod dens tilfældige og materialistiske eller ikke-åndelige natur).
Jeg kan ikke undgå at bemærke, at Darwins arbejde om evolution synes at være et udemærket eksempel på den videnskabelige metode som Popper senere foreslog. Darwin opsøgte allerede i første udgave de problemer han kunne få øje på, og brugte dem til at afprøve sine teorier, og han reagerede på kritik, ikke ved at ignorere den, men ved at inddrage den i senere udgaver og metodisk overveje dem. Denne åbenhed gør, at det stadig er en fornøjelse at læse hans bøger.
(Jeg vil ikke her gå ind på hvorfor og hvordan Darwin kom frem til sine ideer, en gennemgang heraf findes fx i Hoffmeyers bog 'Naturen i hovedet'.)
Det er nu på tide at opridse den mest accepterede forklaring på evolutionen. Den kaldes neodarwinisme eller den moderne syntese, fordi den tager udgangspunkt i Darwins teori og den senere genetiske viden.
Den fælles betegnelse til trods, er der ikke noget der ligner enighed blandt neodarwinister, et mere eller mindre fælles grundlag lader sig dog tegne.
Darwins originale påstand var, at naturlig udvælgelse var den mest betydende mekanisme i evolutionen og ikke mindst, - at den kunne føre til dannelsen af nye arter.
Begrebet naturlig udvælgelse indeholder (mindst) tre forudsætninger:
(1) Indenfor en art er individerne forskellige.
(2) Forskellene er (i hvert fald delvis) arvelige.
(3) Individerne får mere afkom end der 'er plads til'.
Begrebet fokuserer på individerne, et fokus der delvist er forladt i den nuværende neodarwinisme.
Naturlig udvælgelse (fravalg havde måske været mere betegnende) er den proces hvorved de overskydende individer sorteres fra, forhindres i succesfuld reproduktion (i at 'genfrembringe' sig selv ved at få afkom).
Ordet udvælgelse antyder en proces med et minimum af tilfældighed, som nutidens husdyravl; og netop tilfældighedens andel i evolutionens gang har været diskuteret meget.
Darwin skriver:
"For alle væsener må der være megen tilfældig aflivelse, som vil have lille eller ingen indflydelse på den naturlige udvælgelses retning ... men af de der overlever, vil de bedst tilpassede have en tilbøjelighed til at udbrede deres slags i større antal end de mindre godt tilpassede."
(Darwin 79, s 78f)
Den del af udvælgelsen der ikke er tilfældig, tilskrives 'tilpasning' eller 'adaptation' af individet i dets miljø, de bedst tilpassede har størst chance for at overleve og reproduceres.
Desværre bruges de samme begreber om resultatet af udvælgelsen (tilpasning måles ofte ved reproduktion), hvad der har ført til den kritik, at naturlig udvælgelse blot er en tautologi, en ringslutning (fx Popper, se afsnit 1.7).
Jeg tror kritikken beror på en uberettiget brug af logik.
I logikken kaldes logiske sammenhænge i et afgrænset system af tanker for en tautologi. Henvisning til en tautologisk sammenhæng indenfor et sådant system giver ikke nogen 'begrundelse', ingen yderligere information.
Hvis den naturlige udvælgelse var led i et sådant tautologisk system var kritikken ganske berettiget, men det er den ikke.
Den naturlige udvælgelse er ganske vist en slags ringslutning, men ikke en logisk ringslutning. Udvælgelsen foregår i tiden, kan man sige, eller - udvælgelsen foregår i en ring med mulighed for tilfældige ændringer. Der findes forskelle over tid, forandringer, i den ring, som den naturlige udvælgelse er et led i. En ring af fortløbende afprøvninger, af forslag til overlevelse og faktisk overlevelse eller ej.
En undersøgelse af sammenhængene omkring naturlig udvælgelse kan derfor give yderligere information, ny information fremkommet ud af kaos, ud af tilfældige forandringer.
'Ringen' indeholder ikke blot information om den faktiske overlevelse eller ej, men også information om forandringernes vekselvirken med overlevelsen.
Det er i denne vekselvirken vi må finde en forståelse af historien, af de faktiske hændelser.
Den naturlige udvælgelse som princip afhænger således af en bestemt analogi til udvælgelsen i avlsarbejde, 'overlevelsesfordelen' eller tilpasningen må være til stede inden udvælgelsen finder sted, uafhængigt af den faktiske overlevelse eller ej.
(Bl.a Gould 78, s 41)
Neodarwinismen antager at naturlig udvælgelse styrer evolutionære forandringer, at den er den skabende eller styrende proces og (imt. Darwin selv), at tilfældighed og andre mekanismer kun indvirker på evolutionens gang i mindre eller ubetydeligt omfang.
Her i ligger bl.a, at de arvelige forskelle må være små og tilfældigt fordelt, hvis forskellene var store ville dannelsen af forskellene være den styrende proces.
(Bl.a Hoffmeyer 84, s 99ff og Gould 80, s 119f)
En konsekvens af disse antagelser, der fastholdes som central af neodarwinister, er at artsdannelse repræsenterer tilpasningsændringer på alle trin, over gradvist forandrede mellemformer.
(Naylor 85)
Jeg kommer nok ikke udenom at forsøge at definere begrebet art her.
Alle naturinteresserede ved, at det som regel lader sig gøre at henføre de individer man iagttager til en bestemt art. Det forunderlige, set i lyset af en stadigt forandrende evolution, er egentlig, at vi finder en sådan orden i individernes mylder. Skyldes dette menneskets vilje til at finde opdelinger, grupper og grænser, for at kunne kommunikere om og reagere på, kontinuerte fænomener; som når smogadvarsler kommer i under middel, middel og over middel? Eller er grænserne mellem arterne mere reelle, som mellem individer, eller er det en mellemting?
I 1940'erne fandt man frem til det biologiske artsbegreb, som står centralt i neodarwinismen:
" En art er et reproduktivt fællesskab af populationer, som er reproduktivt isoleret fra andre og som optager en specifik niche i naturen."
(Her fra Mayr 82, s 273ff)
To ting falder i øjnene her; der tales ikke om individer men om populationer, fordi reproduktionen er det centrale; men den økologiske rolle inddrages også.
Populationen er et vigtigt begreb, fordi populationen eksisterer indenfor vores egen tidshorisont. Går vi ud og foretager økologiske undersøgelser i naturen, finder vi at individerne hænger sammen via slægtskab. Ofte kan vi finde en mere eller mindre veldefineret grænse, over hvilken vi ikke finder reproduktive forbindelser, denne grænse afgrænser en population, en basal enhed i økologien.
Økologisk niche er en anden mulig enhed i naturen, hvor opdelingerne er foretaget efter, så at sige, den adfærd der ikke har med reproduktion og andre rent sociale forhold at gøre, altså populationens berøringsflade med det økosystem den indgår i.
Arten føles som et mere abstrakt begreb, hvis vi skal definere den indenfor populationens tidshorisont, hvilket jeg egentlig ikke tror kan lade sig gøre.
Vanskelighederne ligger i at definitionen må bygge på potentielle forbindelser mellem populationer.
Afgørelsen af om der består et reproduktivt fællesskab mellem to populationer eller ej, lader sig ikke afgøre ved eksperimenter, men kun ved (minimalt indgribende) iagttagelse.
'Ren' iagttagelse findes ganske vist ikke; som før nævnt vil iagttagelse altid være en selektiv proces, vi må vælge at iagttage en del af helheden. Men i praksis er det muligt at betragte komplekse biologiske fænomener, som reproduktion, på en måde der forstyrrer meget lidt.
Til gengæld vil enhver større manipulation med individer eller miljø betyde, at vi ikke kan overføre resultaterne ukritisk på forholdene som de var inden vi greb ind. Vi finder fx mange 'artskrydsninger' i zoologiske haver.
Jeg mener dette problem hører til den gruppe af fænomener, hvor videnskaben støder på noget den hverken kan bevise eller modbevise. Eller formuleret på en anden måde; fænomener, hvor de reduktioner, vi kan foretage, ikke giver de svar vi søger.
Problemet er beslægtet med det, Tor Nørretranders har behandlet i 'Viden og ikke-viden', hvor emnet er forskning omkring kræftfremkaldende stoffer. Vi kan ikke iagttage de faktiske forhold, og må derfor iagttage en afvigende situation, oftest i form af et eksperiment.
Fastlæggelsen af en bestemt iagttagelsessituation, hvor visse forhold manipuleres, fører til en ikke-viden om de ikke-iagttagne faktiske forhold, der er lige så relevant for problemet som den opnåede viden, men som ofte (bevidst eller ubevidst) ignoreres.
De reduktioner vi foretager i disse to situationer er i begge tilfælde nødvendige reduktioner, som vi ikke kan sætte os ud over, det er reduktioner i tidsdimensionen.
Kræftfremkaldende stoffer bør tilbageholdes eller fjernes inden de fremkalder kræft hos mennesker, mens tidshorisonten for en iagttagelse af populationernes sammenhæng ligger udenfor vores direkte rækkevidde.
Problemet er så, at vi må indse, at de svar vi får ikke er dækkende; at undersøgelsen har givet os en ikke-viden, som er relevant for vores opfattelse af fænomenet.
(Nørretranders 87)
3.3 Flere slags arter:
Morfologiske (i form eller udseende) og fysiologiske ligheder er tidligere blevet brugt som (det eneste) grundlag for henføring til den ene eller anden art. Men sådanne arter er af og til blevet opdelt i to eller flere, eller flere arter er blevet slået sammen til en, efter studier af individernes adfærd og relationer (fx fisken Blåstak og Rødnæb, der ikke har nogen fælles artsbetegnelse på dansk; Blåstak er de gamle hanner).
Inddragelsen af en 'specifik niche' i artsbegrebet, er et forsøg på at udvide begrebets gyldighed.
Artsbegrebet blev formuleret af forskere med erfaringer hovedsageligt fra populationer af pattedyr og fugle. Men reproduktion er et langt mere forskelligartet fænomen end disse populationer giver indtryk af. Mere eller mindre aseksuel (ukønnet) vegetativ formering (fx deling, knopskydning, rodudløbere) er udbredt i andre dyregrupper, og hos planter, svampe og mikroorganismer. Der er da også stor forvirring omkring artsbegrebet hos sådanne organismer.
Niche-kriteriet skulle fx muliggøre henførelsen af separate ukønnede kloner til samme art. Men det fører ofte til modsigelser; nogle mikroorganismer (fx de der lever i afgrøder) har fx vidt forskellige økologiske nicher, men henføres alligevel til samme art.
Muligvis er det håbløst at forsøge at definere en enhed, der skal kunne tjene til opdeling af alle slags levende væsener.
Tilbage står, som jeg var inde på under populationen, at der er flere indgange til at indele naturen i enheder. Slægtsfællesskab eller reproduktion er en mulighed, berøringsflade med ('funktion' i) økosystemerne en anden, morfologi en tredje.
Disse indgange fører ikke nødvendigvis til de samme enheder, og hvis de gør det for én type organismer, er det ikke ensbetydende med at de gør det for andre typer.
Begreber som arten er ikke lige anvendelige overalt i den organiske verden. Bruges begreberne (alligevel) ukritisk, fører det ofte til forvirring og manglende forståelse.
Lignende overvejelser kan gøres over artsbegrebets rolle set i et større tidsperspektiv. Da artsbegrebet indgår i næsten al argumentation om evolution, er det uhyre vigtigt at gøre sig klart hvilke grænser der er for dets anvendelse. Og jeg tror artsbegrebet (eller flere lignende begreber) er vigtigt, på trods af de ovennævnte problemer; jeg vil i det følgende vende tilbage til det.
Jeg vil nu se på et par af de punkter, der kan kritiseres i den moderne syntese. (Jeg mener som mange andre kritikere at den neodarwinistiske teori ikke er helt forkert, men helt utilstrækkelig.)
Evolutionsteori har i lang tid fokuseret kraftigt på genetiske aspekter, specielt populationsgenetik, dvs. fordeling og overførsel af DNA (der bærer det meste af den arvelige information) gennem generationerne i en population.
En nyttig betegnelse for udvikling af populationsgenetisk type er fylogenese, udvikling af slægtskabslinier.
Til gengæld er ontogenesen, udviklingen fra æg til voksent individ (og død), gledet ud af fokus. Evolutionen er oftest blevet set som en evolution af voksne individer, gennem naturlig udvælgelse af voksne individer.
Francois Jacob udtrykker det således:
"Evolutionen beskrives i fylogenetiske udtryk, det vil sige som dannelse af forskelle mellem voksne organismer. Men forskellene mellem disse voksne organismer afspejler altid kun forskelle mellem de udviklingsprocesser, som frembringer disse voksne. Det er især gennem et helt net af kontrolinstanser over for individudviklingen, den naturlige udvælgelse virker, idet den filtrerer de fænotyper, der realiseres udfra de mulige genotyper. For virkelig at forstå evolutionsprocessen må man først forstå fosterudviklingen. Først da vil man kunne bedømme omfanget af de forandringer, der er forenelige med en organismes organisationsplan og funktionsmåde, og bestemme evolutionsspillets regler og betingelser. Desværre ved man endnu næsten intet om fosterets udvikling."
(Jacob 85, s 89f)
Ontogenesens mekanismer er efter min mening nutidens bedste bud på et mysterium. Selvom vi bruger nogle årtier på at kortlægge menneskets arvemasse, vil vi stadig befinde os meget langt fra en forståelse af, hvordan denne arvemasse, det befrugtede ægs øvrige bestanddele, og en vis mængde information og energi fra omverdenen bliver til et menneske.
Når jeg nævner information fra omverdenen i denne forbindelse, er det dels fordi information udefra vides at fastlægge symmetriakser, som op/ned og frem/bag, i ægget.
(Bl.a Løvtrup i Bonde 87, s 97ff)
Men der er også en anden grund. Biologen Rupert Sheldrake har for nyligt lanceret en teori om formdannende felter, som endnu ikke er blevet tilstrækkeligt afprøvet, og derfor heller ikke afvist. (Men den blev faktisk afvist af mange ledende forskere lige på stedet. Den blev bl.a kaldt 'den bedste kandidat til bogbrænding i mange år' af tidsskriftet Nature's, redaktør, hvilket vel nærmest taler til gunst for bogen og til ugunst for redaktøren. - Det var før Rushdie.)
Det vigtigste spørgsmål bogen stiller er: Må vi i udgangspunktet for form- og mønsterdannelse af enhver art medtage tidligere form og mønsterdannelser?
Sheldrake beskriver hvordan former, der tidligere har eksisteret i naturen, synes at påvirke de som senere opstår således, at formerne gennem gentagelse bliver stadig mere stabile:
"Formdannelsesfeltet fra et tidligere system bliver nærværende for alle senere, lignende systemer med en kumulativ virkning tværs gennem rummet og tiden. Når en form - for eksempel et trekløver, først en gang er opstået, vil den efterlade sig et abstrakt billede i et bagvedliggende immaterielt plan, og dette 'formbillede' øger chancen for at en tilsvarende kløver skal opstå igen. Hver gang det sker, bliver billedet skarpere og bidrager stadigt mere til at sikre gentagelser af en stabil form. Princippet gælder alt fra atomer og molekyler til planter og dyr, og virkningen svækkes altså ikke med tid og afstand."
(Her fra Dammann 87, s 324ff)
Nu er der nok nogen, der bliver forskrækket over sådanne immaterielle påvirkninger, men det synes jeg ikke der er grund til.
Dels minder dette meget om den informationsverden jeg tidligere har været inde på. Dels er de 'materielle' påvirkninger i vores verden forårsaget af såkaldte felter, fx tyngdefelter og elektromagnetiske felter, der ganske vist udbreder sig i tid og rum, men ellers ikke er spor 'materielle'.
Opfattelsen af visse dele af virkeligheden som mere materielle end andre knytter sig til vores erfaringer med ting i vores egen størrelsesorden; i den kvantemekaniske verden har begrebet ikke samme relevans.
Fuglene har orienteret sig i jordens magnetiske felt, længe før vi, gennem grundige undersøgelser og tilvænning, fik bragt disse påvirkninger ud af det magiske og overnaturlige område og ind i den selvfølgelige og naturlige dagligdag.
Med hensyn til uafhængigheden af tid og rum, har jeg jo tidligere været inde på Aspect-eksperimentet, og fysikeren David Bohm har også før det arbejdet med en helhedsteori, som bl.a indebærer forbindelser udenfor tid og rum.
(Bohm 86)
'Det er jo umuligt'-argumentet har derfor mistet meget af sin tidligere vægt, og den videnskabelige prøvbarhed må, også i dette tilfælde, være afgørende for teoriens accept.
Princippet om formdannende felter gælder for form- og mønsterdannelse generelt, ikke bare i ontogenesen; og de mest lovende områder for prøvning af teorien ligger tilsyneladende indenfor indlæring og krystalliseringer af nye stoffer.
De foreløbige afprøvninger har givet nogen støtte til teorien, og jeg mener den har så interessante implikationer, at den fortjener bedre end at blive ignoreret.
(Sheldrake 81, her fra Dammann 87, s 324ff)
"Man kan ikke tro på noget, der er umuligt," sagde Alice. "Så har du ikke øvet dig ret meget," sagde dronningen. "... jeg har somme tider troet på hele seks umulige ting inden morgenmaden."
(Lewis Carroll: Bag spejlet)
De genetiske videnskaber har fejret store triumfer på mange områder, men ukendskabet til ontogenese udgør et problem for forståelsen af evolutionen.
Der er et svælg mellem vores viden om overførsel, ændring og blanding af arvematerialet DNA, og vores viden om individernes adfærd, konkurrence, overlevelse og død. Vi ved en masse om hvordan instruktionsbogen ser ud men næsten intet om hvordan cellerne skaber in-form-ation ud af den, hvordan formerne skabes udfra bogen. Og da disse former udgør grundlaget for individernes livsforløb, og dermed for den evolutionære proces, synes svælget at befinde sig lige midt i den evolutionære syntese, neodarwinismen.
Jeg vil senere berøre denne problemstilling igen.
Spørgsmålet om, hvorvidt det levende og vores jord (eller for så vidt hele universet) har gennemgået en evolution, kan vi kun besvare ved at ekstrapolere udfra de begrænsede erfaringer, vi kan opnå gennem vore korte livsforløb.
Vi oplever alle forandringer i vores liv, og også irreversible forandringer. I den biologiske verden kan vi iagttage forandringer, dyrearter uddør, husdyr forandres og på nogle områder kan vi også finde forandringer i naturlige populationer.
Det er dog ikke helt simpelt at konkludere her udfra, at der på jorden er sket en biologisk evolution. Der opstår problemer når vi undersøger fænomener med en tidshorisont, der er langt større eller mindre end vores egen. Vi er da afhængige af en eller anden form for ekstrapolation, dvs. prøve at danne billeder af begivenheder i andre tidshorisonter udfra vores egen.
(Med tidshorisont mener jeg ikke kun tidsskalaens udstrækning, men også dens enheder, processernes 'tidskorn'.)
Ved sådanne metoder har vi dannet os et billede af universets historie tilbage til et punkt for omkring 10-15 mia. år siden, kaldet The Big Bang. Her går grænsen for denne (vores eneste?) metodes muligheder. Vi siger at universet blev skabt i dette punkt, fordi vi ikke kan vide noget udover denne grænse.
Når vi ekstrapolerer, taler om hændelser uden for det område vi kan undersøge eksperimentelt, må vi forudsætte at visse regler gælder udenfor den del af tid og rum vi har erfaring fra.
Det er almindeligt at forudsætte, at de regler der anses som uimodsagte indenfor vores erfaringsområde, er 'universelle og evige' regler. Dvs. at reglerne, der ofte kaldes naturlove, er gældende for det univers vi har mulighed for at gætte om, fra dets tilbliven til det evt. forsvinder igen (heriblandt reglerne for rum og tid!?).
Reglerne kan enten være opstået sammen med universet, være iboende i vores univers, eller nogle kan være udefrakommende i en eller anden forstand. Jeg nærmer mig her til et meget generelt og vanskeligt spørgsmål, - hvor findes de regler vi kan påvise for et vist system, og hvordan foregår 'effektueringen' af reglerne?
Dette spørgsmål stilles i et helt nyt lys af det førnævnte Aspect-eksperiment, der ikke kunne afvise en sammenhæng udenfor vores normale rum og tid.
Jeg vil forlade spørgsmålet og vende tilbage til jordklodens evolution.
Et spørgsmål, der kan åbne den undersøgende tanke, er: Hvad udvikles? Der er sket forandringer, men forandringer af hvad? Former, funktioner, samspil, relationer, eller mønstre? Gener, organismer, populationer, arter? Fødenet, økosystemer, biosfæren eller er det jordkloden som et ubrydeligt hele?
Her dukker problematikken omkring helhed og del, analyse og syntese op igen. Sådanne spørgsmål kan ikke besvares med et enten-eller; men kan bruges til at finde komplementære svar, alternative opdelinger og indgange til et vanskeligt emne, hvor ingen enkelt indgang kan give den maksimale forståelse.
Der er mange indgange til en undersøgelse af evolutionen.
Geologiske undersøgelser giver et slags (mangelfuldt) aftryk af visse former og tilstande (fx fossiler og kontinentaldrift) til forskellige tider, der gennem en fortolkning kan give et billede af jordens historie.
Sammenligninger og klassificeringer af organismer og deres bestanddele, kan ved hjælp af andre fortolkninger give et billede af en anden type.
Økologiske analyser, adfærdsiagttagelser og udviklingsanalogier fra andre historiske forløb, er også blandt de mulige indgange jeg kan se.
Men de billeder, der kan fremkaldes gennem de ovennævnte indgange, indeholder alle elementer af ekstrapolation; og de dertil knyttede problemer, som jeg har været inde på ovenfor, er derfor centrale i en kritik og udvikling af fremkaldeprocessen, fortolkningen.
En sådan kritik og diskussion har stået på i flere århundreder. Jeg vil her blot give et enkelt eksempel.
"Natura non facit saltum", skal Carl von Linné have sagt i det 18. århundrede, - naturen foretager ikke spring.
Linnés 'spring' var dog spring i den opstigende rækkefølge man kaldte naturens stige, huller i guds skaberværk.
Diskussionen for og imod gradualisme (evolution uden spring) har stået på længe, og (måske delvist på grund af frygt for eller håb om associationer til samfundsmæssige revolutioner) været ret ophedet.
"Jeg forstår De er tilbøjelig til at støtte muligheden af betydelige spring i Moder Naturs variationer. Jeg har altid haft den samme opfattelse, til Hr. Darwins store misfornøjelse."
Sådan skrev Thomas Huxley til William Bateson (der for øvrigt er Gregory Batesons far), allerede i 1894.
I diskussionen ligger bl.a spørgsmålet om hvilke regler fra mikroevolutionen (nutidig eller eksperimentel evolution), der er gyldige i makroevolutionære (udenfor vores tidshorisont) fortolkninger, og måske specielt hvilke regler der derudover har spillet ind.
Dette spørgsmål vil jeg senere overveje nærmere.
(Bl.a Gould 77a, s 115)
Kapitel 4 Indhold
DET HIERARKISKE ASPEKT
Når vi vil undersøge en organisme kan vi reducere den fysisk, splitte den op i mindre dele og undersøge disse hver for sig. Dette vil give os viden om bestanddele og strukturer i organismen.
Men vi kan også forsøge at reducere organismens eksistens på en anden måde, vi kan betragte organismen som en helhed, et system. Dette system vil udvise en eller anden form for 'adfærd'; eller rettere adfærden er vores begrundelse for at se på netop organismen som et system. Et system kan kun opfattes som en helhed gennem dets interaktioners dynamik. Det dynamiske og det systemiske er samme ting.
Den systemiske reduktion er netop en reduktion af den samlede adfærd til interaktioner eller relationer. Dette kan give os viden om hvordan systemets bestanddele fungerer som en helhed.
Det er her systemernes hierarkiske aspekt viser sig. Vi kan undersøge delsystemer i organismen, fx organer eller enkelte celler som systemer i sig selv. Og vi kan undersøge organismen som et delsystem i et større system, fx en familie, et økosystem eller en population. Det er denne 'system i system' opdeling, der her kaldes hierarkisk; altså ikke som en rangorden i en hønsegård, men mere som en mængde kinesiske æsker.
At betragte naturen og dele af naturen som systemer vil altså bl.a sige at være opmærksom på sådanne egenskaber, der er egenskaber ved hele systemet og ikke ved dets dele. Disse egenskaber kan repræsenteres af et sæt af variable, der netop vælges udfra, at de antages at belyse relevante egenskaber. Systemet må altid undersøges ved hjælp af et udvalgt sæt af de mulige variable, hvis vi skal have mulighed for at give vores viden videre.
Et eksempel er de undersøgelser af vores jordklode som et system, der i øjeblikket foretages, hvor de variable er temperatur, kuldioxidindhold i atmosfæren og i havet, ozonindholdet i den øvre atmosfære mv.
Vi må også her være opmærksom på, at de valg vi foretager af systemer, hierarkiske niveauer og variable netop er valg; de er mere eller mindre vilkårlige, og der vil (imt. de kinesiske æsker) oftest være alternative muligheder.
Jeg vil senere komme nærmere ind på systemegenskaber i forbindelse med evolution, men her vil jeg først se på kommunikation igen, og derefter på regulering i individet.
4.2 Logiske typer, klasser og paradokser:
Bateson definerede information som en forskel, der gør en forskel. Med udgangspunkt i denne grundlæggende enhed i sansning, vil jeg forsøge at vise nogle hierarkiske aspekter i sansning og kommunikation.
Vi har været inde på, at enhver billeddannelse indebærer en fortolkning eller transformation; en transformation af de forskelle der er blevet opfattet eller udvalgte.
Bateson skriver om forskelle:
"... der er forskel på forskelle. Hver effektiv forskel betegner en skelnen, en klassifikationslinie (at vi ser en forskel indebærer at vi har opdelt det sete i to forskellige slags, to forskellige klasser), og al klassifikation er hierarkisk. Med andre ord forskellene selv må adskilles og henføres til klasser."
(Oversat fra Bateson 85, s457)
Forskelle i tekstur er forskellige fra forskelle i farve, forskelle i størrelse er forskellige fra forskelle i form osv.; og disse forskelle mellem forskelle er igen forskellige.
Bateson siger at al inddeling i klasser må være hierarkisk, at klasserne ikke befinder sig på samme niveau. Han henviser her til en metamatematisk (om matematik) teori, udarbejdet af A.N. Whitehead og Bertrand Russel i 'Principia Mathematica', kaldet 'teorien om logiske typer'.
Teorien handler om forholdet mellem en klasse og dens elementer, og dens centrale påstand er, at en klasse ikke kan være element i sig selv og, at et element ikke kan være hele klassen.
Udtryk der bruges om klassen er af et andet abstraktionsniveau, en anden logisk type, end de udtryk der bruges om elementerne.
Teorien blev udviklet for at undgå de paradokser, der er følgen af kategorifejltagelser, af at forveksle klassen med dens elementer. Paradokser var almindelige i logikken og matematikken på den tid og forekommer overalt i mindre formel kommunikation.
Et eksempel vil vise at fænomenet er velkendt og i princippet ligetil:
Du siger, at hunden er hvid med sorte pletter og din søster siger, at hunden er et ældgammelt husdyr. Hvis lillebror så skriver i sin stil at hunden er et husdyr med sorte pletter, har han taget fejl af kategorierne hund og hund, af klassen af hunde eller hundearten og elementet Trofast. Hvis han ikke indser sin fejltagelse, kan den føre til paradokser, problemer i hans forståelse af hunden og hundearten.
Andre kategorifejltagelser kan føre til (eller er) sygelige tilstande.
Brugen og erkendelsen af sådanne fejltagelser opfattes i visse situationer som humor, der udgives fornøjelige bøger med fejltagelser som lillebrors.
Et klassisk eksempel på et paradoks er kretenseren Epimenedes udsagn: "Alle kretensere lyver".
Det udgjorde et paradoks for mange filosoffer, fordi de ikke klart skelnede mellem kretenseren (elementet) Epimenedes og klassen af kretensere. Hvis han taler sandt, så lyver han og hvis han lyver, så må han tale sandt.
Som filosofferne måske ville have forstået, hvis ikke fremmede folkeslag var så mistænkelige, var det kun Epimenedes der løj. Kretenserne som klasse havde ikke kun valget mellem at lyve eller tale sandt, der var også den mulighed at nogle talte sandt og nogle løj - som Epimenedes.
4.3 Logiske typer og kommunikation:
Begrundelsen for at bruge teorien om logiske typer udenfor matematikken ligger i dens nytteværdi. Kommunikation i den 'virkelige' verden lader sig delvist analysere på samme måde som den del af matematikken der kaldes logikken.
Blot må vi huske at være opmærksom på de punkter hvor analogien ikke gælder fuldt ud, dvs. den ikke-viden vi skaber ved at bruge logikken som model for verden. Som jeg var inde på under omtalen af naturlig udvælgelse i afsnit 3.1, adskiller logikkens ’årsagssammenhænge’ sig bl.a fra vores beskrivelse af virkelige årsagssammenhænge ved ikke at indeholde tid.
Selvom teorien om logiske typer kan være klargørende for vores snak om verden, må vi altså være opmærksom på grænserne for dens meningsfulde anvendelse. Kommunikation udgør ikke hele verden (men hele vores diskussion om verden!), og kun på dele af kommunikationens verden kan vi direkte anvende teorien i den her beskrevne udformning.
Kommunikation består af en udsendelse af potentiel information, der kan være målrettet i større eller mindre grad, og en opfangning og omformning, hvorigennem informationen bliver til og kommunikationen forbinder afsender og modtager.
Omformning af information kan ske på to principielt forskellige måder.
Omformningen kan være analog, dvs. relativt ensartet eller gradvis, hvorved den information der dannes er analog til eller ligner den oprindelige information. Eller omformningen kan være digital, dvs. med spring eller tærskler; afhængig af springenes 'størrelse' og udformning vil den dannede information have mindre eller ingen umiddelbar lighed med det oprindelige. Digital omformning kan fremtræde helt som analog, som en CD'er lyder som en LP.
Oftest vil kommunikation bestå af en blanding af begge dele.
Eksempler kan være det analoge fotografi af en skovsø, og den digitale beskrivelse der gives i et stykke tekst; en mundtlig beskrivelse vil indeholde begge elementer, fx med (mest) digitale ord og (mest) analoge armbevægelser og stemmeføring.
Teorien om logiske typer (i den her nævnte form) gælder kun for digital kommunikation, idet der ikke findes negation (nægtelse, ikke ...) i analog kommunikation.
Bateson stræbte efter at lave en lignende teori for ikke-digital kommunikation.
(Bateson 85, s 279ff og 457f og Ølgaard 86, s 54ff og 111f)
Et uddrag fra Batesons metalog (dialog, hvis opbygning genspejler dialogens emne) 'What is an instinct?' handler om negation i adfærd:
Datter: ... Er der noget 'ikke' i dyrs adfærd?
Far: Hvordan det?
D: Jeg mener, kan et dyr sige gennem dets handlinger, 'Jeg vil ikke bide dig'?
F: Hmm, for det første. Kommunikation med handlinger kan ikke have tider (fx datid, ønskemåde). De er kun mulige i sprog. ...
D: OK. Men hvad med 'ikke'. Kan dyret sige 'Jeg bider dig ikke'?
F: Der er stadigvæk en tid i det. Men det er lige meget. Hvis dyret ikke bider det andet, så bider det det ikke, og det er det.
D: Men det kan være mange andre slags ting dyret ikke gør; sove, spise, løbe osv. Hvordan siger det 'Det er at bide, jeg ikke gør'?
F: Det kan det kun, hvis bidning på en eller anden måde har været nævnt.
D: Mener du, at det kunne sige, 'Jeg bider dig ikke' ved først at vise tænder og så lade være med at bide?
F: Ja. Noget i den retning.
D: Men hvad med to dyr? De ville være nødt til at vise tænder begge to.
F: Ja.
D: Og så, forekommer det mig, kan de let misforstå hinanden og komme i slagsmål.
F: Ja. Der er altid den fare når du handler i modsætninger, og ikke kan eller vil sige hvad du gør, specielt når du ikke ved hvad du gør.
D: Men dyrene ville vide, at de viste tænder for at sige, 'Jeg vil ikke bide dig'.
F: Jeg tvivler på, at de ville vide det. I hvert fald ville de ikke vide det om den anden. ...
D: Så er det en slags eksperiment - .
F: Ja.
D: Så de kommer måske i slagsmål for at finde ud af, om det var 'at kæmpe' de skulle.
F: Ja - men jeg vil hellere sige det mindre overlagt - at slagsmålet viser dem hvilken slags forhold de har, bagefter. Det er ikke planlagt.
D: Så 'ikke' er der faktisk ikke når dyrene viser tænder?
F: Det tror jeg ikke . Eller ikke ret tit. Gamle venner kan måske slås for sjov, og vide fra starten hvad de gør.
(Oversat fra Bateson 85, s 54f)
4.4 Kommunikation og kontekst:
Vores digitale, analytiske kommunikation om verden må ifølge de ovennævnte betragtninger være hierarkisk i sin natur; men Bateson argumenterer for at sådanne betragtninger er nyttige overalt hvor emiske aspekter spiller en rolle, hvor sansning indgår i hændelsesforløbet, - altså også hvor kommunikationen er mere eller mindre analog.
Et argument for nytten af logiske typer knytter sig til den fortolkning eller omformning, der indgår i processer som sansning, og som jeg tidligere har været inde på.
Hvis en organisme skal fortolke en vis mængde forskelle som information, fx et sammenhængende budskab, kræves det at den forud for eller i forbindelse med budskabet har erhvervet sig information om, hvordan forskellene kan hænge sammen og hvordan de skal fortolkes. (Bemærk analogien til den kvantemekaniske iagttagelsessituation i afsnit 2.6.)
Information om fortolkning eller omformning af potentiel information er af en anden logisk type end den information der fås ved fortolkningen. Konteksten, situationen eller sammenhængen indvirker på, eller er med til at definere, hvordan potentiel information opfattes eller fortolkes.
Et eksempel på dette er, at vi ikke kan afgøre om et skub i en flok drenge er en provokation til slagsmål eller en opfordring til leg, hvis vi ikke er i stand til at få information om konteksten. (Relationen, forholdet mellem individerne, udgør en vigtig del af denne og lignende kontekster, som leg, kamp, reproduktion og omsorg.) Denne information kan fx ligge i tidligere erfaringer med individerne, eller et eller andet sted i den potentielle information hændelsesforløbet indeholder.
Begreber som årsag og virkning kan kun bruges entydigt om fænomener, der er afgrænset til ét logisk niveau. Konteksten kan ikke møde et budskab på lige fod, fx bekræfte eller modsige det, men man kan sige at konteksten klassificerer forskellene, henfører forskellene til klasser, til den ene eller den anden slags. Hvis du genkender ham der skubber, kan dette give dig information om hvordan skubbet skal fortolkes, om det skal i klassen 'provokation' eller i klassen 'lege-opfordring', men det kan ikke modsige skubbet.
En Ockhams ragekniv (et sparsomhedsprincip) gælder her som for andre forklaringer; man bør ikke forøge de logiske typer unødvendigt.
(Bateson 84, s 109ff og Ølgård 86, s 58ff)
Bateson fører denne argumentation langt videre i en omfattende model af indlæring og mind. Jeg vil stoppe med disse indicier for sansningens og kommunikationens hierarkiske natur, og se lidt på kommunikation eller regulering i en organisme.
Jeg har tidligere berørt genetikkens vigtige rolle i neodarwinismen. Genetikken har, siden James D. Watson og Francis Crick fik æren for opdagelsen af DNA's rumlige struktur i 1953, været en af de hurtigst voksende videnskaber - i penge, omfang og betydning. Der går næppe en dag uden at genetiske emner, specielt gensplejsning, kommer frem i medierne og debatten.
Generelt er opfattelsen af grundlaget for arvelig overføring og ontogenetisk udvikling præget af et fokus på en bestemt del af substansen, DNA, eller måske snarere den information som DNA-molekylerne bærer. DNA betragtes som en slags instruktionsbog, der på en eller anden måde læses i cellen.
Dette er sandsynligvis et rimeligt billede, men de overvejelser genetikerne (forhåbentlig) har om hvordan forandringer i genotypen (individet karakteriseret ved DNA) bliver til forandringer i fænotypen (individets fremtrædelsesform som organisme) finder sjældent ud i debatten.
Det er almindeligt at forstå denne omsætning, omtrent som en computerstyret industrirobot, der læser i et program og omsætter dette til en fremstillingsproces. Det er sådanne opfattelser, der ligger bag neodarwinismens overseelse af ontogenesen, som jeg har omtalt i afsnit 3.4.
Betragtninger om information, omformning, regulering og logiske typer er nødvendige, hvis vi skal kunne vurdere, hvorvidt computeranalogien er berettiget eller ej.
(Se fx Hoffmeyer 88 og Fjerdingstad 88, for et indlæg i debatten om genteknologi, der inddrager sådanne betragtninger - og et svar, der repræsenterer den manglende forståelse for de samme betragtninger.)
Jeg har ikke brugt begrebet 'gen', fordi der i begrebet ligger den opfattelse, at genomet (et individs 'sæt' af DNA) er opdelt i afgrænsede områder, hver ansvarlige for et 'arveanlæg'. Denne enhed i genomet har kun en begrænset anvendelighed i genetikken.
Det er almindeligt at bruge betegnelsen gen om et område på DNA-molekylet, der bærer information om sekvensen i en enkelt proteinkæde. Begrebet gen hænger således sammen med, at en stor del af den molekylærgenetiske forskning foregår på prokaryoter (fx bakterier og blågrønalger) og virus. Hos prokaryoter findes nemlig ofte en sådan entydig sammenhæng.
Men prokaryoter er langt simplere organismer end eukaryoter (fx gær, amøber, svampe og højere dyr og planter). Og encellede eukaryoter er igen simplere end de flercellede, hvis liv udviser et ekstra niveau, i organiseringen af cellerne under ontogenesen.
Regulering af transskriptionen (kopiering af DNA til RNA) og translationen (oversættelse fra RNA til proteinkæde) hos prokaryoter er kendt i detaljer.
Reguleringen hos encellede eukaryoter som gær, alger og amøber, minder på flere områder om prokaryoternes, pga. lignende livsvilkår, men organisationen af genomet og cellen er som hos de flercellede eukaryoter.
Hos disse findes der ikke nogen simpel sammenhæng mellem DNA og proteinkæder, deres genom har en langt mere kompleks organisation. Det DNA, der koder for en proteinkæde, kan fx ligge spredt på genomet, og er næsten altid fyldt med ikke-kodende sekvenser (introns).
Hos flercellede eukaryoter, som planter, insekter, bløddyr, fisk og firbenede dyr og fugle, er reguleringen af ontogenesen som hele næsten ukendt, dog er reguleringen af enkelte proteiner delvist kendt, specielt blandt de der produceres i meget stort omfang.
Kommunikationen i flercellede organismer varetages delvist af specialiserede langstrakte celler der indgår i det vi kalder nervesystemet. Overførslen af information i nervesystemet sker dels gennem pulser af elektriske spændingsforskelle i nervetrådene, og dels gennem overførsel af kemiske stoffer (transmitterstoffer), hvor nervecellerne er i kontakt med hinanden (i synapserne). Kemiske stoffer indgår også i reguleringen af celler udenfor nervesystemet, disse stoffer kaldes hormoner (nogle transmitterstoffer er også hormoner). Hormonerne kan produceres bestemte steder i organismen og derfra transporteres til andre steder, hvor de kan indvirke på bestemte målceller. Målcellerne besidder nogle receptorer (modtagere), der formodes at bære hormonet gennem cellekernens membran, og måske medvirke i reguleringen. Herudover ved man meget lidt, bortset fra at transskriptionen på en eller anden måde sættes i gang af hormonet. Det vides ikke hvordan syntesen af receptorer begrænses til målcellerne.
Hormoner vides at spille en rolle i ontogenesen, fx styres sommerfuglenes forpupning og metamorfose (forvandling) af mængden af et bestemt 'ungdoms'-hormon.
Hormonproduktionen kan variere i et ontogenetisk bestemt mønster, men er ofte påvirkelig af ændringer i det ydre og indre miljø.
(Hoffmeyer 84, s 126f og Snyder 85, s 392-428)
Det fremgår af det ovenstående, at vores viden om den genetiske regulering i flercellede organismer er langt fra udtømmende på nuværende tidspunkt. Denne indgang alene kan altså ikke give os den forståelse af ontogenesens rolle, som vi søger.
De generaliseringer af matematik og logik jeg hidtil har været inde på, og andre generelle betragtninger om systemer kan muligvis bidrage til en sådan forståelse.
4.6 Regulering i ontogenesen:
Vores ideer om reguleringen må kunne redegøre for de i naturen observerede ontogenetiske forløb.
Hos mennesker og lignende dyr er det ontogenetiske forløb tilsyneladende gradvist fremadskridende. Spørgsmålene her er bl.a, hvordan cellerne differentierer (bliver forskellige). Hvordan disse forskelle bliver til i en organiseret sammenhæng, hvordan forskellene organiseres til et komplekst hele.
Og endvidere i et populationsperspektiv; hvordan disse helheder holdes tilstrækkeligt ensartede til at tiltale det andet køn og muliggøre overleven i og med miljøet, men alligevel tillades at være tilstrækkelig forskellige, så de kan danne grundlag for en evolution.
Hos nogle organismer må reguleringen redegøre for spørgsmål af en anden type; nemlig det fænomen at forskellige fænotyper på skift kan udfolde sig fra det samme genom, fra den samme genotype.
Eksemplerne er almindelige og forunderlige; frøer og insekter (fx sommerfugle) udfører iøjnefaldende forvandlinger som de fleste har set med egne øjne. Planters varierende vækstformer, forskellen på kaster blandt sociale insekter, kønsforskelle og skift af køn er andre almindelige eksempler.
Akimoto har studeret arter af rodlus (slægtninge til bladlus), der skifter mellem to værtsplanter i løbet af et år, og udviser op til tre helt forskellige fænotyper.
Et eksempel er ribsrodlusen, der overvintrer på elmetræer. De første (jomfrufødende) generationer på elmetræerne lever i galler på bladene; de sidste får vinger og flyver til planter af ribsslægten, hvor generationer af en anden fænotype trives. Igen får den sidste generation vinger og flyver tilbage til elmetræerne, hvor en kønnet larvelignende generation fødes, parrer sig, lægger æg og dør.
(Jørgensen 78, s 23)
Akimoto skriver, at der kun har været få forsøg på at inkorporere sådanne fænomener i evolutionsteori, eller på at klarlægge mekanismerne. Nogle skift i fænotype er helt uafhængige af ydre påvirkninger, mens økologiske faktorer spiller ind i andre tilfælde.
En evolutionsteori, hvis gyldighed skal strække sig udover pattedyr og fugle og gælde for alle organismer, må inddrage konsekvenser af rodlusenes livshistorier og lignende fænomener.
Sådanne fænomener er indicier for det nødvendige i at bruge hierarkiske forklaringsmåder om genomets organisation og regulering, og eksempler på, som Akimoto siger det:
"Genomers enorme fænotypiske potentiel som en materiel basis i makroevolutionen."
(Akimoto 85)
En vigtig konsekvens af disse betragtninger er, at forandringer i genomet ikke kan betragtes som ligeværdige. Nogle genotypiske forandringer må være af en anden logisk type end andre.
Forandringer i genomet, mutationer eller arvelige forskelle, er grundlaget for neodarwinismens naturlige udvælgelse. Men ontogenesen ligger mellem de arvelige forskelle, og de individer der er objekter for den naturlige udvælgelse. Når tilsyneladende ens forskelle i genomet udfolder sig til at være forskelle af vidt forskellig type hos individerne, kommer den underliggende antagelse i neodarwinismen til syne; at oversættelsen og 'udfoldelsen' af genotypen er ensartet, og at forholdet mellem genotype og fænotype basalt er 'en til en'. Denne antagelse holder ikke.
Jeg vil i det næste kapitel se nærmere på individernes ontogenese i sammenhæng med populationen. Men det er måske nyttigt først at se på udvikling på en lidt mere generel måde, end jeg har gjort i forbindelse med ontogenesen.
4.7 Nogle typer af proces eller udvikling:
Et forløb i tid, en udvikling eller proces, kan indeholde elementer af kaos, tilfældighed og nødvendighed.
Det nye, emergens, kreation eller uforudsigelighed er knyttet til, ja betinget af kaos og tilfældighed. Processer der er uforudsigelige, ikke bare i praksis men i princip, kaldes divergente. Modsvarende kaldes forudsigelige processer (der kun indeholder uforudsigelighed i et omfang eller af en type der ikke influerer på de forudsigelser vi ønsker at gøre) for konvergente.
Hvis vi klassificerer de forskelle over tid, forandringer, processen består af kommer Russells logiske typer til at spille ind. Beskrivelsen af den divergente proces må indeholde flere logiske typer.
De 'tilfældige forandringer' kan således opdeles i bestemmelige eller forudsigelige klasser, muligheder eller udfald, og de uforudsigelige elementer heri. Indenfor en enkelt klasse, må forskellene kaldes kaotiske.
Begrebet tilfældighed betegner det forhold der findes mellem flere klasser med kaotiske elementer. Eksempler på kaotiske elementer i en enkelt klasse er brownske bevægelser af støvpartikler (pga. sammenstød med luftmolekyler); eller det stjerneformede brudmønster der opstår når et projektil rammer vinkelret på en homogen glasflade med en vis hastighed.
Bemærk at hvis eksempelvis støvkornene ikke er lige store, hvis glasfladen ikke er helt homogen, eller hvis indfaldsvinklen eller hastigheden af projektilet varierer, er der mulighed for opdeling af de kaotiske elementer i flere klasser. Er denne opdeling fornuftig og rimelig, opnår vi en øget grad af forudsigelighed. Det tilfældige adskiller sig fra det kaotiske ved dets indhold af mønster, og mønsteret er grundlaget for vore forudsigelser.
Et eksempel på en proces med flere forskellige udfald eller klasser er punktmutationer, forandring på enkelte positioner i DNA-kæden. Forandringer i 'syntaks (sætningsbygning) og stavning' kan opdeles i klasserne udskiftning, tilføjelse eller fjernelse af 'bogstaver' (nukleotider). Endvidere kan forandringerne, for hver position, klassificeres efter hvilke konsekvenser de har for transskription og translation.
Genomet indeholder information af flere typer, såsom information om aminosyrer, om aminosyrernes sammenkædning til proteinkæder og om forholdet mellem (reguleringen af) oversættelsen af forskellige proteinkæder.
Desuden er der en vis redundans (overflødighed) i kodningen af ovennævnte information. På visse positioner vil nogle forandringer i syntaks og stavning derfor have en langt mere drastisk effekt på transskription og translation end andre.
Endelig må der, som jeg har nævnt, i et ontogenetisk perspektiv være forskellige typer af forandringer af genotypen. Forandringer der virker ind på forskellig måde i den overordnede regulering af organismen og dens udvikling.
Tilfældighed er som sagt af en anden type end både elementerne og klasserne; tilfældighed betegner en relation eller et forhold mellem klasserne.
Hvis dette forhold er relativt konstant (hvis processen forløber under bestemte betingelser), kan det beskrives med statistiske begreber, som sandsynlighed, udfra visse antagelser om tilfældighedens fordeling.
Statistikken er nyttig derved, at den kan demonstrere og belyse forhold, der er langt mere komplekse end de her anvendte. Men når statistik bruges til at afgøre om et resultat skal anses for at 'kunne være tilfældigt' eller at 'sikkert være ikke-tilfældigt', så indgår de statistiske antagelser (om fordeling mv.) blandt de antagelser, der ligger til grund for fortolkningen. Statistikkens anvendelighed afhænger altså af gyldigheden af dens antagelser.
(Bl.a Kragh 81, s 44ff)
Vi vil kalde en proces divergent, hvis vore forudsigelser omfatter de kaotiske elementer (der udspringer af det grundliggende kaos som kvantemekanikken viser os), mens den samme proces kan kaldes konvergent, hvis vore forudsigelser gælder elementer af andre logiske typer, hvor det kaotiske har lille eller ingen betydning. Undersøgelser af et enkelt kuldioxidmolekyles fastlægning i fotosyntesen belyser en divergent proces, mens undersøgelser af den samlede fotosyntese omfatter forudsigelser af andre typer, og derfor belyser processen som en mere forudsigelig og konvergent proces.
Hvis en divergent proces foregår i et system der indeholder evnen til at sammenligne de i processen fremkomne forskelle, kaldes den en stokastisk proces. Ordet er græsk og betyder at skyde med bue mod et mål; skudfordelingen er i begyndelsen ret kaotisk, men bueskytten, buen, pilene og målet ændrer på dette ved at sammenligne forholdene omkring hvert skud.
Man kan se det som en divergent proces, der bliver overlejret af en konvergent proces, idet sammenligningen betyder en øget forudsigelighed eller bestemthed.
Evolution i neodarwinisternes formulering er et eksempel på en stokastisk proces, hvor sammenligningen kaldes naturlig udvælgelse. Andre processer, der kan betragtes som store stokastiske processer, er indlæring og immunitet.
(Inspireret af Bateson 84, s 31ff og 187f)
Kapitel 5 Indhold
INDIVID OG POPULATION
Jeg vil nu, inspireret af Bateson, nærme mig mikroevolutionens problemer på en deduktiv måde, dvs. jeg vil formulere spørgsmål og problemer omkring individ og population udfra nogle af de grundlæggende betragtninger, jeg har berørt i de foregående kapitler.
5.1 Forandring i individ og population:
Forandring og stabilitet må altid defineres ved henvisning til at en eller anden beskrivelse vedbliver at være sand eller ej, så deres type fastlægges.
En linedanser opretholder en stabil balance ved stadige forandringer i forholdene mellem kroppens dele. Eller skrækken bevirker en dag et stabilt forhold mellem kroppens dele - og balancen forandres.
Homøostase er betegnelsen for linedanseren i vores krop. Kroppens indre miljø indeholder et netværk af tilbagevendende kontrolløkker (feedback løkker), den selvkorrigerende balance gælder fx temperatur, saltbalance og pH. Lignende homøostaser findes mange andre steder i den levende verden, ilt- og kuldioxidindholdet i atmosfæren, temperaturen i biosfæren og saltindholdet i oceanerne er stabile tilstande i den homøostase vores jordklode har udvist i årmillioner, indtil nu. Ofte er de stabile tilstande lette at beskrive, mens de korrigerende forandringer, sammenvævede som de er, er sværere at rede ud.
(Bateson 84, s 57f og Lovelock 87)
I biologien findes der ikke variable der udelukkende stiger eller falder i tiden, de har alle et eller flere tolerable områder, og minimum og maksimum herfor.
Som Paracelsus sagde i det 16. årh.:
"Was ist das nit gifft ist: Alle ding sind gifft und nichts ohn gifft - allein die dosis macht das ein ding kein gifft ist."
Men de tolerable områder varierer for forskellige organismer.
Der findes således store forskelle i udstrækningen af de forskellige arters homøostase.
Et eksempel, der ofte drages frem, er temperaturreguleringen - vekselvarme og ensvarme dyr.
Man kan klassificere arterne efter, i hvilken grad de holder visse interne variable konstante, til trods for ændringer i de tilsvarende eksterne miljøvariable. Dette gælder miljøvariable som temperatur, relativ luftfugtighed eller mere generelt tilgængeligheden af vand, saltindhold eller osmotisk tryk i vandige miljøer, iltindhold, kuldioxidindhold og pH, og koncentrationen af forskellige andre kemiske stoffer.
Alle organismer opretholder en vis homøostase, visse biologiske variable holdes ret konstante eller organismen dør. Men de variable, der svarer til miljøets variable, holdes ikke konstante i samme grad eller udstrækning for forskellige slags organismer.
Grænsen for variablernes homøostase kan ligge indenfor, på, eller udenfor organismens grænse. Vekselvarme dyr tillader temperaturen i kroppen at falde i visse perioder (dette gælder også nogle pattedyr og fugle). Andre dyr og fugle vedligeholder en homøostatisk temperatur udenfor kroppen, fx redebyggende dyr og fugle, sociale insekter m.fl.
Begrænser vi os (foreløbigt) til flercellede eukaryoter, kan forandringer i individer og populationer ifølge Bateson klassificeres i tre hovedtyper:
Jeg har først et par bemærkninger til denne klassificering.
Ontogenesens forandringer må antages i princippet at kunne deles i to slags: Dels en 'tautologisk' del, udfoldelsen af genotypen uden indvirken 'udefra'; denne del vil jeg (for at undgå forvirring) i det følgende kalde epigenese (udvikling der bygger på det forudgående). Og dels en 'tilfældig' eller variabel del, de somatiske forandringer.
Tidshorisonten for de forskellige forandringer er ikke den samme, dette skal senere vise sig at være af stor betydning.
Batesons klassifikation kan bl.a bruges til at klarlægge argumentationen omkring arvelighed og nedarvningens mekanismer.
5.2 Arvelighed og reproduktion:
Celler må betragtes som de grundliggende materielle og energetiske enheder i det levende (jeg betragter virus og deslige som 'løsrevne fragmenter', der ved interaktion med celler kan kopieres og mangfoldiggøres). Slægtskabslinier forbindes af celledelinger; dvs. en vækst - en kopiering af mønstre og forøgelse af mængder, efterfulgt af en deling, der sikrer at cellens karakteristiske mønstre findes i begge datterceller. Disse celledelinger kaldes mitotiske delinger eller blot mitose.
Biologisk arvelighed er et andet udtryk for den kopiering af mønstre, der finder sted i reproduktionen.
Oftest indgår der udover mitosen, mere eller mindre regelmæssigt, en hel eller delvis sammensmeltning af celler i reproduktionen, dette kaldes seksuel eller kønnet formering.
Hos prokaryoter kan der ske en overførsel af dele af genomet i en proces, der kaldes konjugation; denne overførsel er ikke obligatorisk.
Hos eukaryoter smelter to celler oftest helt sammen til én, eller der overføres et helt genom. Derfor er den 'befrugtede' celle (zygoten) diploid. Dvs. den har to sæt kromosomer (hos eukaryoter er genomet opdelt i kromosomer), et fra hver celle, og i øvrigt strukturer og organeller mere eller (meget) mindre ligeligt fra begge celler.
Inden zygoten (eller dens diploide datterceller) på ny kan smelte sammen med en anden celle, må den igennem en reduktionsdeling, kaldet meiotisk deling eller meiose.
Under meiosen udveksler hvert kromosompar homologe stykker af kromosomerne gennem overkrydsning, normalt mindst en overkrydsning per kromosompar. De to sæt kromosomer fordeles tilfældigt i den efterfølgende deling, så der bliver et sæt i hver dattercelle, der altså igen er haploid.
(Bl.a Snyder 85, s 31ff)
Hos nogle organismer forbliver cellerne efter mitosen i mere eller mindre nær kontakt og fungerer som et hele, som en flercellet organisme eller en koloni.
Oftest er det kun visse celler i den flercellede organisme, kønscellerne, der deltager i den kønnede formering, resten af organismen kaldes soma. Soma kan være dannet gennem mitoser af zygoten, og altså være diploid (fx pattedyr), den kan være haploid (mange alger), eller der kan være både diploide og haploide somaer (fx mosser). Endelig kan nogle individer være diploide og andre haploide; fx hos de sociale insekter som myrer og bier, hvor de haploide individer er hanner.
(Faktisk kan denne struktur i formeringen have været afgørende i netop disse insekters udvikling af komplekse samfund, idet hunner af disse insekter deler en større arv med deres søstre end med mødre og fædre.)
(Se fx Gould 83b, s 58ff.)
Opdelingen i hanner og hunner, eller mere generelt i dannelsen af store immobile kønsceller (æg) og små mobile kønsceller (sædceller eller pollen) er vidt udbredt. Det findes fx hos alle landlevende dyr og planter.
De resulterende zygoter kan herved blive uhyre store; blommen i et strudseæg er således en enkelt celle.
Evnen til at formere sig ukønnet ved simpel deling i to findes kun blandt nogle simple flercellede organismer som søanemoner og fladorm. Knopskydning eller vegetativ formering findes hos mange organismer, fx planter. Sporedannelse er en anden udbredt metode til ukønnet formering.
Ofte vil både kønnet og ukønnet formering forekomme i organismernes livsforløb.
Hos mere komplekse flercellede organismer, som fx insekter og hvirveldyr, forekommer ukønnet formering kun i form af en tilbagedannelse af den kønnede formering til en ukønnet; dette kaldes det jomfrufødsel eller partenogenese.
Sådanne jomfrufødsler adskiller sig fra den oprindelige ukønnede formering ved, at dattercellerne dannes gennem en meiotisk deling.
(Bl.a Gamlin and Vines 87, s 45ff og 235ff)
5.3 Kønnet formering og meiosens betydning:
Tilstedeværelsen af kønnet formering i store dele af den levende verden leder til spørgsmålene om, hvad der ligger til grund for en sådan organisering af reproduktionen eller, - hvilke implikationer den har.
Det egentlige grundlag for en organismes eksistens i en længere tidshorisont er overlevelse 'udover' sig selv, ikke på grund af aldring men pga. den tilfældige udslettelse; i vores verden kan overlevelse kun ske ved at skabe kopier.
Mens en bakterie, der deler sig, overlever fuldt ud (næsten) i begge datterceller, vil en encellet alge kun have en halv andel i zygotens datterceller. Hvordan kan denne tilsyneladende ulempe opstå og udbredes?
I økologiske undersøgelser finder vi, at ukønnet formering typisk findes hos koloniserende eller 'lev mens du gør det' organismer, der lever i nyopståede eller periodiske miljøer. Her er kønnet formering øjensynlig ikke nogen fordel.
Det har alligevel ofte været fremført, at den kønnede formerings fordel er bedre tilpasningsmuligheder, pga. den genetiske variation, der skabes gennem kombineringen af genotyper.
Men denne fordel giver ingen forklaring på, hvorfor evnen til ukønnet formering er forsvundet.
Desuden viser nyere matematiske analyser, at kun meget specielle selektionsmønstre, der er sjældne i naturen, kan vedligeholde kønnet formering i en population, der også kan formere sig ukønnet.
Jeg var i forrige afsnit inde på, at når ukønnet formering findes hos komplekse flercellede organismer, sker det altid gennem en meiose (undtagen for de allersimpleste); dette er et indicium for meiosens betydning for reproduktionen af komplekse organismer.
Der findes mekanismer i cellerne, der kan reparere punktmutationer i DNA-kæden; det er kun de mutationer, der undslipper denne mekanisme, der kan spille en rolle i individernes ontogenese.
Overkrydsningen i meiosen muliggør kombinering af nyttige mutationer eller, sagt på en anden måde, udskillelsen af skadelige. En bestemt organisme vil altså kunne fremme sin egen reproduktion gennem kønnet formering, ved at få en mulighed for at indgå i en succesfuld kombination.
Det er nok denne fordel, der har spillet den største rolle i udviklingen af kønnet formering. Og den er gyldig for både prokaryoter og eukaryoter.
Men spørgsmålet om hvorfor kønnet formering er blevet enerådende kræver et andet svar.
Lia Ettinger foreslår, at meiosens funktion, hos organismer med genomer over en vis kompleksitet, er at vedligeholde genomets grundliggende organisation.
Eukaryoternes genom består for en stor del af 'overflødig' DNA; heriblandt nogle gentagne DNA-stykker, der kaldes repetitiv (gentaget) eller selvisk DNA. Selvisk, fordi det reproducerer sig selv i en hurtigere takt end resten af genomet; der bliver altså mere af det.
I denne DNA-reproduktion spiller de såkaldte transposomer (springende gener) en vigtig rolle. Transposomer virker også i dannelsen af kopier af gener, og medvirker dermed til den mulighed for større genetiske spring sådanne kopier af gener udgør (fordi de er ’ledige’ til forandring).
Den dynamiske og flydende karakter, som eukaryoternes genom udviser, er således en kilde til evolutionære nyskabelser.
Men på den anden side må der være grænser for DNA-reproduktionen; dels fordi den er en trussel mod den basale organisation, og dels fordi det er en omkostning for organismen at slæbe rundt på alt for store genomer fyldt med overflødig DNA.
Det er her meiosen bevirker en bortselektering af celler, hvor der er sket ændringer i den basale organisation.
Fordelen for organismen ved denne selektion er dels, at den kan udvælge de kønsceller, der sandsynligvis er funktionsdygtige, og dermed have større muligheder for at danne fungerende zygoter. Dette vil specielt være en fordel for organismer der kun kan reproducere sig gennem relativt få zygoter, som pattedyr, fugle og mange insekter.
Men selektionen holder også konstant styr på det selviske DNA; uden en sådan kontrol ville genomet blive større og større, og udgøre en større og større byrde for organismen.
Selve udvælgelsen sker sandsynligvis i den fase i meiosen hvor de homologe kromosomer ligger parret i hele deres længde og overkrydsningen sker. Sammenligningen af de to kromosomer sker formodentlig ved hjælp af specielle DNA-sekvenser, der ligger imellem generne på kromosomerne med bestemte mellemrum; da der må være spillerum for forskellige udgaver af generne.
(Det er også uforenelighed i meiosen der gør, at artskrydsninger som muldyr oftest er sterile.)
Den kønnede formerings fordel er altså snarere den ukønnede formerings ulempe; den ukønnede formering uden meiosis findes ikke hos de mere komplekse dyr, fordi meiosen er et nødvendigt led i deres reproduktion.
Lia Ettinger nævner flere undersøgelser, der støtter hendes teori: Undersøgelser af nematoder viser, at de somatiske celler har et langt større antal transpositioner (flytten rundt) i deres DNA end kønscellerne. Og en stor andel af kønscellerne hos mange forskellige organismer bliver faktisk elimineret af degenerative processer inden de færdigdannes. (Bl.a Ettinger 86, s 17ff)
5.4 Somatiske forandringer og adaptation:
Forandringer i miljøet, både det levende og det ikke-levende, er et uundgåeligt fænomen på vores klode, også indenfor enkelte organismers livsforløb.
Organismerne kan overleve sådanne forandringer på forskellige måder. De kan måske tolerere dem, hvis forandringerne ikke influerer på dem. De kan tilpasse sig midlertidigt, overfor kortere og måske regelmæssigt tilbagevendende forandringer. Eller de kan tilpasse sig mere permanent.
Den enkelte organismes tilpasninger sker ved somatiske ændringer, dens muligheder herfor betegner organismens fleksibilitet. Jeg vil herefter kalde denne type tilpasning for adaptation. Ikke alle somatiske forandringer er adaptive.
Det ovennævnte må ses på baggrund af kroppens homøostase af sammenvævede variable, hver med begrænsede muligheder for forandring, som jeg har beskrevet tidligere.
En konsekvens heraf er at en somatisk forandring har indflydelse på den samlede fleksibilitet. Hvis forandringen betyder at en eller flere variable nærmer sig deres grænser, vil forandringer der ændrer disse variable i samme retning bevirke en overskridelse af grænserne, og organismen bliver 'skadet', den dør måske. Fleksibiliteten kan genoprettes ved at andre forandringer ændrer variablerne i modsat retning, eller ændrer grænserne for dem - en forandring af grænserne er af en anden logisk type end forandring af variablen selv.
Bateson bruger et eksempel med en kvinde, der bevæger sig fra havoverfladen op på et bjerg (ikke Himmelbjerget).
Forandringen i ilttryk betyder at kvinden må trække vejret hurtigere og hendes hjerte banker hurtigere. Disse umiddelbare somatiske adaptationer sænker hendes fleksibilitet betydeligt; hun vil fx ikke kunne yde samme fysiske arbejde som hun kunne ved foden af bjerget.
Men hun er bjergbestiger og venter derfor midt på bjerget for at akklimatisere sig. Akklimatiseringen indebærer mange forandringer, blodets indhold af hæmoglobin øges, hjerte- og brystmuskulaturen ændres osv.; men forandringerne sker ikke lige hurtigt og er ikke lige reversible.
Kroppen bruger fleksibilitet på et 'dybere' niveau, og genvinder herved den umiddelbare fleksibilitet. Eller man kan sige at den fordeler belastningen fra enkelte kritiske variable på flere eller mindre følsomme variable; måske gennem en ændring af nogle af variablernes grænser.
Organismens overlevelse bygger altså på opretholdelsen af en øjeblikkelig og generel (og derfor kostbar, nødvendig i mange situationer) fleksibilitet. Dette sker gennem adaptationer af en mere specialiseret og langsomt omsættelig type; adaptationer til de miljøforandringer, der viser sig at være mere permanente.
Udstrækker forandringens permanens sig til populationens tidshorisont opstår muligheden for, at den somatiske adaptation mere eller mindre kan erstattes af tilpasning gennem genotypiske forandringer.
5.5 Genotypiske forandringer og tilpasning:
Der er vigtige forskelle på karakteren af somatiske og genotypiske forandringer.
Men generelt ved vi for lidt til i praksis at skelne mellem en epigenetisk udfoldning af en genotypisk forandring i ontogenesen, og en somatisk forandring der giver samme fremtrædelsesform.
Der ligger en væsentlig forskel i hvordan forandringen opretholdes i individet.
En somatisk forandring sker som nævnt i et selvkorrigerende netværk af informationskredsløb, og opretholdes i og af dette; den er derfor oftest mere eller mindre reversibel.
Den genotypiske ændring er derimod fastlagt i epigenesen, den er indkodet i hver enkelt celle. Dette betyder dels, at organismen 'sparer' at adaptere sig og opretholde ændringen i det homøostatiske informationskredsløb, men det betyder også, at den oprindelige somatiske tilstand måske ikke længere kan nås, der er måske ingen vej tilbage.
Til gengæld må 'besparelsen' give organismerne en større somatisk fleksibilitet.
Mere generelt kan man inddele de genotypiske forandringer i flere typer, udfra deres indvirken på den somatiske fleksibilitet på forskellige niveauer.
Genotypiske ændringer, som iagttageren er opmærksom på pga. deres 'primære' effekt, vil ofte hvad enten de anses for tilpasninger, neutrale eller skadelige drage veksler på den somatiske fleksibilitet; idet en ændring ét sted (den primære effekt) vil indvirke mange andre steder i det homøostatiske netværk.
Der må derfor også være genotypiske ændringer, der øger organismens fleksibilitet; ellers ville genotypiske tilpasninger føre til et tab af fleksibilitet, og slægtskabslinierne uundgåeligt uddø, som følge af organismernes 'stivnen'.
Hos komplekse flercellede organismer må det endda for hver genotypisk tilpasning til ydre forhold være nødvendigt med mange genotypiske tilpasninger til den indre homøostase.
Disse ændringer er næppe af en sådan art, at de tiltrækker iagttagerens opmærksomhed, men deres eksistens må indirekte kunne vises gennem eksperimenter med en meget kraftig kunstig selektion i en bestemt retning, hvorved fleksibiliteten efterhånden vil gå tabt.
Der er dog åbenlyse vanskeligheder i sådanne eksperimenter; det vil være svært at skelne genotypiske forandringer fra de somatiske adaptationer der sker til dem; og ligeledes vil det være svært at skelne de genotypiske ændringer der giver øget fleksibilitet, fra de 'dybe' somatiske adaptationer de 'aflaster'.
Endelig vil det i selektionseksperimenter være uhyre svært at skelne mellem mangel på fleksibilitet i individerne og mangel på variation i populationen. Hos fx planter, der både kan formeres vegetativt fra et enkelt individ og kønnet i en population, vil denne sidste vanskelighed måske kunne undgås.
Hvis vi ser dette i et evolutionært perspektiv kan vi på den ene side sige, at individets genotype udgør en ramme for, hvilke somatiske forandringer der er tilladelige.
På den anden side må man sige, at mere vedvarende relationer mellem individerne i en population og deres miljø, relationer der er betinget af og udtrykt i individernes somatiske adaptationer, danner en ramme eller en støbeform for de genotypiske forandringer.
Ser vi på populationen, må den somatiske fleksibilitet også udgøre den stødpude, der muliggør eksistensen af populationens genotypiske fleksibilitet.
En population indeholder en stor mængde genetisk variation, der konstant skabes og kombineres på nye måder via den kønnede formering og overkrydsningen, og de (mindre hyppige) kromosommutationer der kan ske under meiosen. De sjældnere punktmutationer i DNA-kæden skaber også genetisk variation i et længere perspektiv.
Den genetiske variation i populationen som helhed udgør populationens grundlag for bl.a at besvare miljøets ændringer. Men de forskellige genotyper kan kun overleve i populationen, og den genetiske variation bevares, i det omfang de somatiske forandringer kan kompensere for konsekvenserne i de enkelte individer.
Men er det rimeligt overhovedet at opdele i somatiske og genotypiske ændringer?
Hos encellede organismer føles denne grænse mellem genotypiske og somatiske ændringer ret vilkårlig.
Genomet udgør blot et led i cellens homøostase, og i mitosen 'arves' både genomet og resten af cellens strukturer og de homøostatisk opretholdte tilstande. Grænsen støttes dog af cellernes udveksling af DNA (hvis det kun er DNA de udveksler?).
Hos de komplekse flercellede organismer indeholder begrebet 'somatiske forandringer' forandringer af en anden logisk type end hos enkelte celler. Den flercellede somas homøostase er af en anden type end cellens; den indeholder et netværk af relationer mellem celler, der er fremkommet gennem epigenesen.
Flercellede organismers somatiske forandringer kan derfor snarere lignes med forandringer i relationerne mellem encellede organismer end med forandringer i en enkelt celle.
Det må (foreløbigt) formodes at epigenesen hos flercellede organismer har sit grundlag i zygoten, udover påvirkninger fra anerkendte felter som tyngdefeltet, og eventuelt en vis nødvendig indvirken fra moderen. Der må derfor være et genetisk (her i betydningen arveligt) grundlag for de somatiske forandringer; det har således ingen mening at spørge om en vis somatisk egenskab (den mest heftige diskussion har gået på indlæring) afspejler miljøets betingelser eller den genetiske arv.
Spørgsmålet må være:
På hvilket logisk niveau spiller det arvelige grundlag ind for denne type forandring?
Visse 'egenskaber' er direkte genetisk fastlagte; et eksempel er øjenfarven, der er kun mindre somatiske forandringer.
På næste niveau er den somatiske forandring fastlagt, det kan fx tænkes at væksten af hår og negle er genetisk bestemt; men det kan også være at det der er fastlagt ikke er væksten, men vækstens relation til kroppens ernæringstilstand mv., dvs. to niveauer fra egenskaben hårlængde.
Ligeledes kunne egenskaben hudfarve tænkes at være fastlagt på dette niveau, således at solbruningen var en vis funktion af sollys mv. for en bestemt pige. Men jeg har hørt nogle sige, at man lettere bliver brun efterhånden, hvis man sørger for at blive brun hvert år. Hvis dette er tilfældet, må det være den mulige ændring i bruningens hastighed, der er genetisk fastlagt, tre niveauer fra hudfarven.
Disse eksempler kunne antyde at arven kunne beskrives ved, at genotypen bestemte differentialkvotienter af stigende orden, men det er nok kun en analogi der gælder for disse simple eksempler.
5.7 Evnen til at skelne:
Helt generelt er det af stor betydning, at der findes processer på flere logiske niveauer og, at de foregår i forskellige tidshorisonter.
At fx genotypiske tilpasninger foregår i en længere tidshorisont end de homøostatiske adaptationer, og derfor næsten altid vil være bagefter (lag behind), kan være en vigtig årsag til, at netop denne type organisation er udbredt.
Det centrale dogme i molekylærgenetikken siger at information kun kan overføres fra DNA til protein, aldrig fra protein til DNA. Og at somatiske forandringer derfor ikke direkte kan indgå i genotypen.
Der er dog ikke noget principielt til hinder for en mekanisme til nedarvning af erhvervede egenskaber hos flercellede organismer. Man ved fx, at proteiner spiller en vigtig rolle i reguleringen af cellernes genom; sådanne proteiner kunne, styret af karakteren af individets ontogenese, overføres til kønscellerne og influere på zygotens epigenese. Eller der kunne tænkes andre mekanismer, som ikke findes i de organismer videnskaben har studeret indtil nu (hvilke apropos overvejende er pattedyr og specielt hvide laboratorierotter).
Det afgørende argument mod en sådan mekanisme er, at flercellede organismer, der besidder en sådan (effektiv) mekanisme, hvis de findes, vil leve i den samme simple tidshorisont som bakterier, fra dag til dag så at sige, med dvaletilstand og kolonisering som de eneste eksistensmuligheder.
Organismernes egenskaber vil blive fikseret i generne 'alt for hurtigt', de vil derfor ikke kunne tolerere forandringer i miljøet gennem somatiske adaptationer.
Evnen til at skelne mellem mere og mindre bestandige forandringer i populationens miljø, og dermed muligheden for en ny type 'livsstrategi', en strategi der baseres på bæredygtighed, ligger netop i eksistensen af kontrol på flere niveauer, over flere tidshorisonter.
Ontogenese og fylogenese består således tilsyneladende af processer der ligner de stokastiske processer jeg omtalte i afsnit 4.7.
Evnen til at skelne er den sammenligning der udgør den konvergente proces' styring af den underliggende divergente proces.
Figuren nedenunder er et skematisk billede af forbindelserne mellem ontogenese og fylogenese, zygote og flercellet soma, og genotype og fænotype.
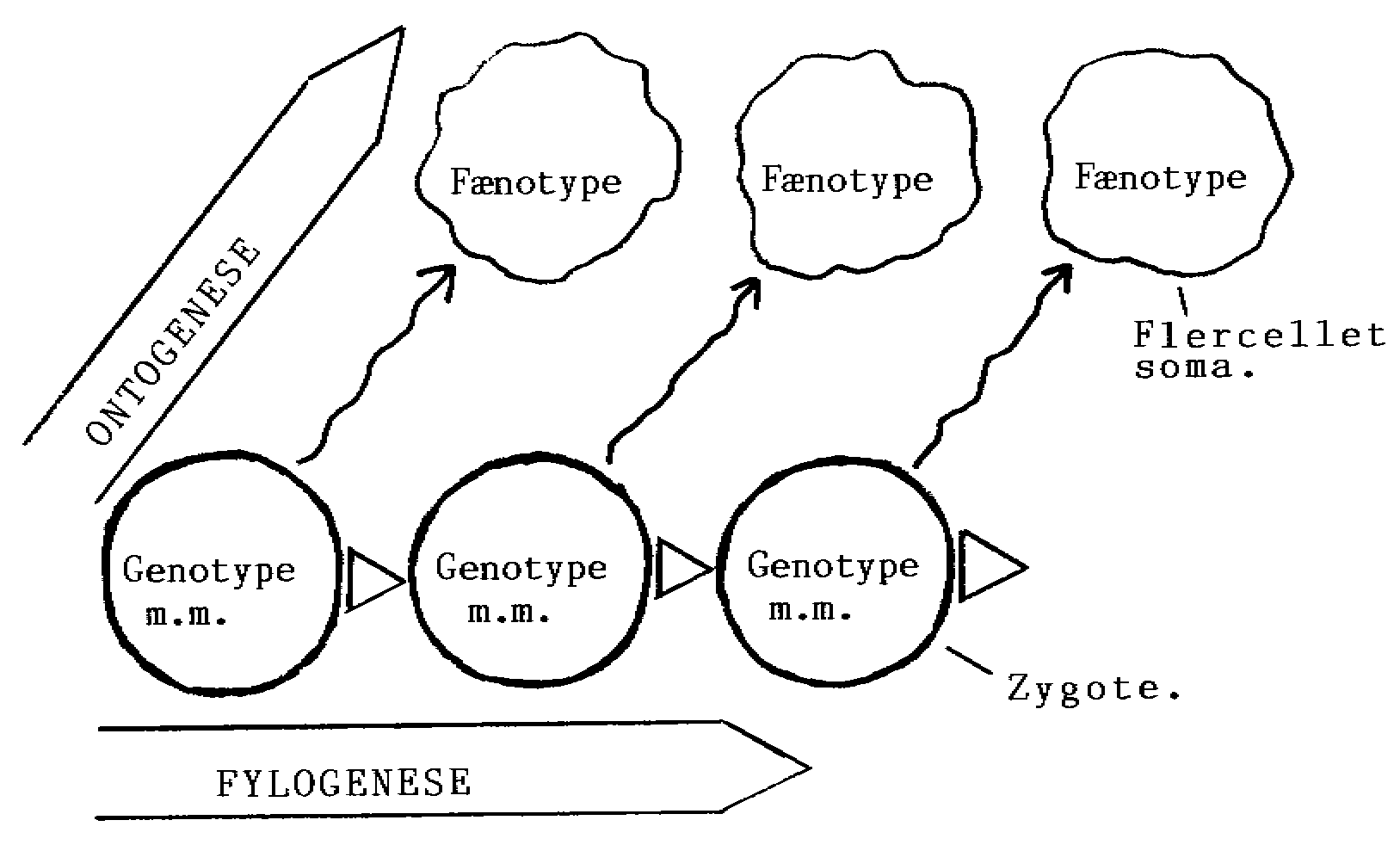
(Ide efter Hoffmeyer 84, s 95)
Kapitel 6 Indhold
AUTOPOIESIS
I dette vil jeg forsøge at samle trådene fra de foregående kapitler, specielt i lyset af de tanker Humberto Maturana og Fransisco Varela lægger frem i 'Kundskabens træ'.
Bogen præsenterer et fascinerende og meget sammenhængende billede af vores erkendelse af verden, med fokus på begrebet 'selvopretholdelse'.
Fordelen ved at bygge på bogens tanker er, at det kan give et nødvendigt overblik over emnerne. Ulempen ligger netop i at Maturanas tanker er så sammenhængende; de repræsenterer et slags paradigme, og udgør derfor en begrænsning overfor alternative tanker.
I et forsøg på at bruge fordelen og undgå ulempen, har jeg valgt at fremstille Maturanas tanker samlet her. Dels som en opsamling, - men samtidig bringer forfatternes indsigt os videre mod en forståelse af den levende verden.
6.1 Erkendelsens cirkulære natur:
Kundskabens træ udtrykker et syn på erkendelse der minder meget om det jeg har antydet i kapitel 2, og altså også om den konstruktive videnskabsteori:
"Der er, som vi også vil komme ind på i det følgende, ingen tvivl om, at vi erfarer en verden. Når vi imidlertid nærmere undersøger, hvordan vi kommer til at erkende denne verden, vil vi uvægerligt finde, at vi ikke kan adskille vores fortids handlinger - biologiske og sociale - fra hvordan verden ser ud for os. Det er så åbenbart og indlysende, at det er meget svært at se."
Videre siger de, at denne cirkulære forbindelse mellem handling og erfaring giver en vis svimmelhed når vi tænker over, hvad det er at vide og erkende; for vores særlige måde at være på og den måde, hvorpå verden ser ud for os er uadskillelige. Enhver videnshandling frembringer en verden, - al gøren er viden og al viden er gøren.
Dette gælder selvfølgelig også for hvad de og jeg skriver;
- alt hvad der siges, siges af nogen.
Enhver tankevirksomhed frembringer en verden, den er en menneskelig handling, udført af en bestemt person på et bestemt sted.
(Maturana 87, s 40ff)
 |
Bogen bygger på begrebet autopoiese.
Et autopoietisk system er et system der er organiseret som et netværk af processer, der producerer de komponenter netværket består af. Man kan også sige, at systemet er cirkulært organiseret og, at det er den cirkulære organisation, der afgrænser systemet. (Se også afsnit 7.3)
Levende væsener udviser en sådan autopoietisk organisation. Heri indgår nogle aspekter, jeg har været inde på.
Homøostasen er identisk med de enkelte organismers autopoiese; det dynamiske netværk af cirkulære interaktioner der opretholder organismen gennem en stadigt pågående selvskabelse ('autos' betyder selv, og 'poiein' at skabe, på græsk).
Det bliver da klart at det netop er denne organismens selvopretholdelse eller autopoiese, der er afgørende for vores opfattelse af organismen som en afgrænset enhed.
Jeg har også berørt organismens regulering, og Maturana foreslår i forlængelse heraf, at autopoiesen er den mekanisme, der gør levende væsener til autonome eller selvbestemmende enheder.
(Maturana 87, s 59ff)
Et uddrag af Gregory Batesons metalog 'Hvorfor har ting omrids?' giver eksempler på denne selvbestemmelse:
Far: Lad os overveje et rigtigt konkret vaskeægte roderi til en forandring, og se om det hjælper. Husker du kroketspillet i 'Alice i eventyrland'?
Datter: Ja - med flamingoer?
F: Det er rigtigt.
D: Og hulepindsvin som kugler?
F: Nej, pindsvin. Det var pindsvin. De har ikke hulepindsvin i England.
D: Åh. Var det i England, far? Det vidste jeg ikke.
F: Selvfølgelig var det i England. Vi har heller ikke hertuginder i Amerika.
D: Men der er da Hertuginden af Windsor, far.
F: Ja, men hun har ikke pigge, ikke som et rigtigt hulepindsvin.
D: Gå videre med Alice og lad være med at være fjollet, far.
F: Ja, vi talte om flamingoer. Pointen er, at manden der skrev Alice tænkte på de samme ting som vi gør. Og han underholdt sig med lille Alice, ved at forestille sig et spil kroket der ville være rent roderi, bare absolut roderi. Så han sagde de skulle bruge flamingoer som kroketkøller, fordi flamingoerne ville bøje halsen så spilleren ikke vidste om køllen overhovedet ville ramme kuglen eller hvordan den ville ramme kuglen.
D: Og lige meget hvordan, kuglen ville måske gå sin vej af sig selv fordi den var et pindsvin.
F: Rigtigt. Så det hele er så rodet at ingen kan sige hvad der vil ske.
D: Og kroketbuerne gik omkring, oven i købet, for de var soldater.
F: Ja - alting kunne bevæge sig og ingen kunne sige hvordan det ville bevæge sig.
D: Skulle alting være levende for at det kunne blive et fuldstændigt roderi.
F: Nej - han kunne have lavet rod ved . . . nej, jeg tror du har ret. Det er interessant. Ja, det var nødt til at være sådan. Vent lidt. Det er underligt men du har ret.
Fordi hvis han havde rodet tingene på hvilken som helst anden måde kunne spillerne have lært at tage højde for roderiet.
Jeg mener, hvis nu kroket plænen var ujævn, eller kuglerne havde en underlig form, eller hovederne på køllerne bare var vakkelvorne i stedet for at være levende, så kunne folkene stadig lære det og spillet ville kun være sværere - det ville ikke være umuligt. Men når man bringer levende ting ind i det, så bliver det umuligt. Det havde jeg ikke ventet.
D: Havde du ikke, far? Jeg havde. Jeg synes det er naturligt.
(Oversat fra Bateson 85, s 30f)
6.3 Information på ny:
Jeg har i opgaven betragtet information og sansning som noget meget betydende i vores forståelse af det levendes væsen, uden egentlig at være istand til at argumentere for det. Maturanas tanker giver mig en mulighed for at knytte en forbindelse mellem det emiske aspekt og det levendes betingelser.
Som følge af den autopoietiske eller homøostatiske forståelse, må (næsten?) alle interaktioner mellem en organisme og dens miljø ligne kommunikatoriske interaktioner.
Maturana udtrykker det på en anden måde:
De ændringer i organismen, som er resultatet af interaktionen mellem det levende væsen og dets omgivelser, kan sættes i gang eller udløses af det forstyrrende medium, men bestemmes af det forstyrrede systems struktur.
(Maturana 87, s 103)
Den strukturelle ændring bestemmes ikke af interaktionen men af organismens foregående tilstand, som er fremkommet igennem dens dynamiske autopoiese og homøostase, interaktionerne er ikke instruktive.
Jeg var selv inde på problemet med at sætte et skel mellem informationsprocesser og 'almindelige' fysiske processer i afsnit 2.4.
Jeg vil nu i forlængelse heraf betragte de fysiske processer som specialtilfælde af informationsprocesserne; de tilfælde hvor vi kan beskrive processen ved entydige årsags/virknings-sammenhænge.
Komplekse systemer, hvor kvantemekaniske processer er af betydning, udvikles igennem historiske processer (se endvidere næste kapitel). Sådanne processer indeholder kaotiske elementer, nogle årsager vil blot være udløsere og nogle virkninger vil blot være påfølgende virkninger, der ikke er bestemt eller fastlagt af årsagen.
Det vil derfor nok være muligt at beskrive og forklare mange informationsprocesser fuldstændigt i fysiske termer.
Men på den anden side tror jeg at alle levende væsener vil udvise interaktioner, som vi formentlig altid må 'nøjes' med at betragte som informationsprocesser.
Kommunikation kan derfor heller ikke betragtes som 'overførsel' af 'information', eller som nogle instruktioner der gives. Kommunikation afhænger ikke af det der overføres, men af det der sker med den, der modtager det overførte.
Informations-, kommunikations- og systemteori indgår blandt vore metoder til at behandle interaktioner med komplekse systemer på en videnskabelig måde.
I disse metoder indgår et skift fra at se på fysiske objekter til at se på relationer mellem fysiske objekter eller, som Bateson siger, forskelle. Begrundelsen herfor må findes i de karakteristika informationsprocesserne udviser. I stedet for præcise kvantitative forudsigelser, må vi nøjes med forudsigelser der drejer sig om klasser og tilfældighed; metoderne må kunne undersøge processer med kaotiske elementer.
En anden begrundelse kan findes i sansningens natur eller 'funktion'; forskelle og forandringer er vigtige for levende væsener i deres livsforløb, mens konstante tilstande ikke er det. Tilvænning i sansningen er sikkert en afgørende mekanisme i organismernes fravalg af det 'for' konstante.
6.4 Autopoiesens arv:
Homøostasen er den enkelte organismes autopoiese, men i Maturanas autopoiese indgår også reproduktionen, som jeg bl.a har behandlet i afsnit 5.2.
Reproduktion betegner helt generelt en kopiering og opdeling, der fører til enheder med samme organisation, altså enheder der henføres til samme klasse som den reproducerede enhed.
De aspekter af den oprindelige struktur i den nye enhed, som vi vurderer som identiske med den oprindelige enheds, kaldes arv; de der vurderes som forskellige kan kaldes reproduktiv variation. Jeg har behandlet sådanne forskelle i en bestemt sammenhæng i forrige kapitel, hvor jeg snakkede om genotypiske og somatiske forandringer.
Som jeg har gjort det, lægger Maturana også vægt på, at arv ikke er begrænset til DNA's domæne, og at DNA nok spiller en væsentlig rolle i arvelighed, men kun som en del af cellens autopoiese eller, med Batesons ord, i den kontekst DNA er en del af.
Et enkelt eksempel på arv der ikke direkte involverer DNA er, at mange strukturer i cellen, som mitochondrier, membraner og støtte- og transportstrukturer, reproducerer sig eller kopieres selvstændigt i cellerne, og at strukturerne hos fx pattedyr altid overføres med ægget i den kønnede formering.
(Mitochondrier og kloroplaster (planternes grønkorn) menes at stamme fra prokaryotiske symbionter eller samlevere, fra meget tidligt i livets historie.)
(Bl.a Maturana 87, s 78ff)
6.5 Strukturel kobling:
Vender vi os mod det autopoietiske systems interaktion med omgivelserne og mere eller mindre lignende systemer heri, kan vi undersøge de gensidige forstyrrelser eller perturbationer (forstyrrelser uden den negative betydning). Interaktionerne må ifølge afsnit 6.3 oftest betragtes som informationsprocesser.
Nogle typer af interaktioner forekommer oftere end andre, denne konstatering har overordentlig omfattende følger. Den konkrete fordeling afhænger af hvilken klassificering vi lægger over interaktionerne.
Mere hyppige eller tilbagevendende interaktioner antyder for os en fastere kobling end de sjældne og enkeltstående interaktioner. Jeg har tidligere været inde på at denne opfattelse ligger dybt i det videnskabelige objektivitetsideal (afsnit 1.8).
Men selvom de tilbagevendende interaktioner ikke siger noget om vigtighed eller betydning, så siger de dog, at vi her har at gøre med koblinger af en speciel type.
Denne type kobling, en historie af tilbagevendende interaktioner, fører (under hvilke omstændigheder?) til strukturel overensstemmelse, og Maturana kalder det for strukturel kobling.
Strukturel kobling er altså en mere generel formulering af de fænomener som jeg hidtil har kaldt adaptation eller tilpasning. En fordel ved denne formulering er at opmærksomheden fastholdes på det gensidige ved sådanne fænomener.
Mit fokus i det forrige kapitel har ligget på de strukturelle ændringer der fører til eller opretholder den strukturelle overensstemmelse. Ændringerne er led i de levende væseners bestandige dynamiske interaktion.
Set i en større tidshorisont overføres eller arves nogle typer af strukturelle ændringer (i afsnit 5.5 har jeg netop set på forholdet mellem de arvelige og de ikke-arvelige ændringer). Karakteren af en bestemt celles strukturelle koblinger må ses som den nuværende tilstand i den række af strukturelle transformationer, der udgør cellens fylogenetiske historie.
Maturana kalder også slægtskabsliniens historie for dens naturlige drift (drift som i 'at drive af sted').
6.6 Metacellulære enheder:
Enhver celle kan betragtes som et autopoietisk og dermed afgrænset system.
I nogle tilfælde synes den strukturelle kobling mellem et antal celler at etablere en særlig ny sammenhæng, der af os erkendes som et større afgrænset system, dette kalder Maturana en metacellulær enhed.
En sådan metacellulær enhed (som fx en gris) udviser et livsforløb af en anden logisk type end den enkelte celle, jeg har været inde på epigenesen som et 'nyt' aspekt i flercellede organismers livsforløb.
En metacellulær enhed kan være en koloni af encellede organismer af samme art, en flercellet organisme eller en symbiotisk enhed med individer af to eller flere arter.
(Det kan være meget vanskeligt at afgøre om der er tale om en koloni eller et individ, nogle gopler som vores egen vandmand regnes således for individer, mens den portugisiske man-o-war udgør en koloni, se fx Gould 85, s 79ff. Også afgrænsningen mellem en symbiotisk enhed og en en- eller flercellet organisme er vanskelig, som fx med de symbiotiske mitochondrier. Sådanne vanskeligheder antyder, at vi forsøger at få en præcis opdeling til at passe med noget, der faktisk mere er et kontinuum.)
Metacellulære enheder der har en autopoietisk organisation, dvs. udviser selvskabelse og selvopretholdelse, kan betragtes som andenordens autopoietiske systemer; og er derfor omfattet af vores overvejelser omkring strukturelle koblinger.
Flercellede organismer udgør et komplekst stadium i ontogenesen af en cellulær udviklingslinie; de indgår (oftest) i en cyklus, i hvilken der er et encellet stadium. Dette er kilde til den overordentligt store strukturelle variation vi kan forundres over, på trods af at den basale organisation er bevaret. Enhver ontogenetisk variation resulterer i forskellige måder at være på i verden, fordi det er enhedens struktur, som bestemmer dens interaktion i miljøet og den verden, den lever i.
(Maturana 87, s 86ff)
6.7 Levende væseners naturlige drift:
Nøglen til en forståelse af evolutionens grundlag - den strukturelle transformations historiske mekanismer, ligger i forbindelsen mellem forskelle og ligheder i den reproduktive rækkefølge. Derfor var Batesons forslag til en sådan forbindelse i forrige kapitel interessant.
Maturanas formulering af dette er både mere generel og mindre anvendelig.
Han opdeler de strukturelle ændringer i en enhed i ødelæggende ændringer, der fører til ændringer i organisation; og tilstandsændringer, der ikke fører til ændringer i organisationen.
Disse ændringer kan enten være udløst af perturbationer fra omgivelserne eller ej.
Organisation betegner i bogen de relationer mellem delene i et system, der er betydende for vores klassificering af systemet, mens struktur betegner de faktiske komponenter og relationer i systemet.
Altså noget der ligner Batesons logiske typer, men begrænset til disse to typer. Netop denne begrænsning gør Maturanas fremstilling lettere forståelig og mindre vidtrækkende end Batesons.
Ser vi nu på begrebet naturlig udvælgelse eller naturlig selektion, er det tydeligt at sproget igen driver sit spil med os.
Darwin pointerede selv, at udtrykket blot var en passende metafor; men metaforer og analogier kan være farlige, senere blev Darwins forbehold glemt.
Naturlig udvælgelse blev opfattet som miljøets 'frie valg' gennem instruktive interaktioner, i mere eller mindre udpræget grad.
Opfattelsen af levende væsener som strukturelt determinerede klargører, at en interaktion ikke kan bestemme, men kun udløse den resulterende ændring. Miljøets 'valg' betegner derfor snarere iagttagerens forståelse af, hvad der sker i enhver organismes livsforløb; udfra sammenlignende iagttagelser af mange organismer.
Den enkelte organisme lever i sin egen historie. De mulige ændringer og 'svar' på miljøets perturbationer for den enkelte organisme er langt fra så mange, som iagttageren tror udfra sine studier af mange organismers historie. I forrige kapitel var jeg mere konkret inde på denne histories, de somatiske ændringers indvirken.
Desuden udtrykker begrebet naturlig udvælgelse som før nævnt en ensidighed, der skjuler den strukturelle koblings gensidige natur.
Levende væsener eksisterer i en ramme der hedder konservering, de er autopoietiske, selvopretholdende systemer. Konserveringen af organisationen sker gennem den strukturelle kobling til organismens omgivelser, den resulterende overensstemmelse kaldes tilpasning af organismen.
Batesons linedanser har fået et nyt kostume på. Organismens historie udviser en strukturel drift i overensstemmelse med miljøets strukturelle drift.
Maturana ser fylogenese og evolutionen som en simpel ekstrapolering af organismernes og miljøets strukturelle drift (stadig drift som i 'at drive af sted'). Organismernes drift i fylogenesen kalder han naturlig drift, en drift langs de forløb, der til enhver tid er mulige, som det fastlægges af organismernes og miljøets historiske forandringer og forholdet imellem dem.
6.8 Teleologi og tilpasning:
Nogle forskere mener at de tilpasninger, vi ser i naturen, er teleologiske eller målrettede, dvs. at de kan forklares ved den funktion de har, ligesom en knivs form er (delvist) teleologisk.
(Fx Fransisco Ayala i Dobzhansky 77, s 497f)
Heri ligger ofte også den opfattelse, at nogle dyr er bedre tilpassede end andre.
Sådanne opfattelser hænger oftest sammen med et fokus på menneskers, pattedyrs eller fugles fortræffeligheder.
Hvis begreber som tilpasning skal have nogen mening, må de anvendes i en sammenhæng, hver art har sin specielle overensstemmelse med miljøet, opstået i en bestemt historisk sammenhæng.
Det er ikke variationerne i miljøet, som en iagttager kan se dem, der bestemmer de forskellige liniers fylogenese.
Fylogenesen bestemmes af det forløb, der er fulgt ved organismernes konservering og strukturelle kobling i deres eget miljø (niche), et forløb, som de definerer, og hvis variationer kan passere ubemærket for en iagttager.
(Maturana 87, s 102ff)
"Med andre ord: da ethvert autopoietisk system er en enhed af mange gensidigt afhængige dele, undergår hele organismen ændringer i mange dimensioner på samme tid, når en dimension i systemet ændres. Men det er klart, at sådanne ændringer, som for os synes relateret til miljøet, ikke opstår på grund af miljøet, men ud af den strukturelle drift, som finder sted i mødet mellem organisme og miljø, der er operationelt uafhængige systemer. Da vi ikke ser alle de faktorer, der er involveret i mødet, synes strukturel drift at være en proces, der er vilkårlig. At dette ikke er tilfældet, skal vi få at se, når vi studerer de måder, på hvilke det sammenhængende hele, som en organisme udgør, undergår strukturelle ændringer."
(Maturana 87, s 120)
| 
|
|
| Far, hvorfor er palmerne så høje? Det er for at girafferne kan spise af dem, mit barn.... | hvis palmerne var helt små, var girafferne i vanskeligheder. | |
| 
|
|
| Men hvorfor har girafferne så lange halse, far? Det er for at de skal kunne spise af palmerne, for... | hvis girafferne havde korte halse, ville de være i endnu større vanskeligheder! |
Det må være rimeligt her at bemærke, at teleologiske forklaringer kan anvendes om visse typer af overensstemmelser mellem organisme og miljø.
Jeg tænker på de dele af eller former for tilpasninger, der svarer til sådanne regelmæssigheder ved verden, som vi beskriver i naturlove; disse regelmæssigheder er så stabile, at de teleologiske forklaringer ikke adskiller sig fra den faktiske strukturelle drift. Det iøjnefaldende eksempel er evnen til at flyve, og det samme eksempel viser, at en meget forskelligartet udvikling af denne evne er forenelig med den strukturelle overensstemmelse, de forskellige arter udviser.
Faren ved de teleologiske forklaringer er, at de er simple og letforståelige, og derfor svære at erstatte med mere korrekte, men oftest også mere komplekse forklaringer. (Se også afsnit 8.2 om bricolage.)
Ifølge Maturana er det altså ikke nødvendigt med en udefra kommende kraft, for at skabe forskellighed og komplementaritet mellem organisme og miljø, ej heller for at forklare en tilsyneladende retningsbestemthed af variationerne.
Evolutionens naturlige drift beror udelukkende på iboende mekanismer.
6.9 Adfærd og viden:
Jeg har ikke i det foregående omtalt adfærd specifikt, til dels fordi jeg ikke ser adfærd som principielt forskellig fra somatiske ændringer; det er vanskeligt at definere en grænse, der skiller adfærd ud fra andre somatiske forandringer.
Begrebet adfærd bruges oftest om forandringer, der foregår i en kort tidshorisont (i forhold til vores egen 'adfærd') dvs. hurtige processer; og endvidere oftest kun om processer, der interagerer med omgivelserne.
(Tilstedeværelsen af et nervesystemet og dets organisation spiller derfor en vigtig rolle i det vi kalder adfærd. Jeg har kort været inde på nervesystemet i afsnit 4.5. Maturana og Varela (der er neurobiologer af fag) giver en dybtgående behandling af nervesystemet, men jeg vil ikke gå ind på det her.)
Med andre ord:
Adfærd er iagttagerens beskrivelse af (det han ser som) tilstandsforandringer i et system, i relation til et miljø som systemet interagerer med; eller mere poetisk - en udenfor ståendes syn på organismens indre relationers dans.
Som følge heraf vil iagttageren ofte beskrive adfærden teleologisk som om det, der bestemmer forløbet af interaktionen, er dens mening eller mål og ikke de interagerende organismers strukturelle koblingers dynamikker.
Erkendelse (forståelse) karakteriseres af Maturana som en effektiv handling, en handling der sætter et levende væsen i stand til at fortsætte dets eksistens i et bestemt miljø, efterhånden som det frembringer sin verden.
Viden må i denne forbindelse ses som iagttagerens opfattelse af organismens erkendelse. Viden er en effektiv (passende) handling eller adfærd i en given sammenhæng, - iagttageren definerer sammenhængen ved de 'spørgsmål', der stilles til organismens viden. Forskellige spørgsmål specificerer forskellig viden, denne forståelse er vigtig i al adfærdsforskning.
Organismer er 'dumme' i en sammenhæng, de ikke passer i, og 'kloge', hvis de er i overensstemmelse med sammenhængen.
(Maturana 87, s 156ff og 192)
En speciel form for strukturelle koblinger er gensidige koblinger mellem organismer af samme art, jeg vil kalde dem sociale koblinger.
Sociale koblinger er en forudsætning for seksuel reproduktion, og dermed en forudantagelse i begreber som population og art.
(Bemærk, at dette er en cirkelslutning, det er vanskeligt at skelne mellem sociale koblinger og strukturelle koblinger med de øvrige omgivelser uden at henvise til arten eller populationen. På den anden side kan man let tænke sig sociale koblinger, der ikke er seksuelle, sådanne koblinger indgår i kolonidannelse hos encellede organismer.
Begrebet sociale koblinger tjener således til at belyse reproduktionen i en større sammenhæng. Man kan måske sige, at sociale koblinger er gensidig hjælp mellem organismer til overlevelse udover den enkelte organisme selv; men kun hvor overlevelsen er 'fælles', og den enkelte organisme også overlever i de øvrige organismers overlevelse.)
I det biologiske artsbegreb ligger altså en forudantagelse om visse sociale fænomener, og gyldigheden af artsbegrebet er derfor begrænset til gyldigheden af denne antagelse; en antagelse, der ikke er nødvendig for organismernes overlevelse. Det var denne begrænsede gyldighed jeg var inde på i afsnit 3.2 og 3.3.
(Se også Maturana 87, s 170ff)
Man skelner konventionelt mellem indlært og instinktiv adfærd. Parallelt med mine bemærkninger i afsnit 5.6 om den genetiske baggrund for somatiske ændringer, må distinktionen mellem indlært og instinktiv adfærd bero på, på hvilket logisk niveau adfærden eller adfærdsmulighederne er arveligt fastlagt; se i øvrigt Bateson, 84.
Adfærd, der ikke er direkte fastlagt igennem arv og epigenese, kan modificeres i løbet af ontogenesen; dette kaldes indlæring.
Indlært adfærd og sociale koblinger skaber i forening muligheden for en kulturel overførsel af adfærd over generationer.
Imitation er den almindelige måde til kulturel bevarelse af interaktionsmønstre (adfærdsformer) hos hvirveldyr.
Kulturel adfærd opstår, bevares og udvikles i sociale systemer gennem imitation og fortsat 'tilpasning'; men den er ikke væsensforskellig fra anden indlært adfærd.
Den adfærd vi kalder kommunikation kan også være instinktiv og/eller indlært. Indlært kommunikativ adfærd kan kaldes sproglig adfærd.
Det følger heraf, at sprog ikke kan betragtes som betegnelser for genstande og fænomener i den 'virkelige' verden, men derimod som ontogenetisk etablerede koordinationer af adfærd.
(Maturana 87, s 182ff)
Jeg har givet dette korte rids af Maturanas argumentation om sociale fænomener, bl.a for at vise hvordan en vis opfattelse af hvorledes naturen fungerer, kan lede til en begrundelse af hvad der ellers betragtes som filosofiske standpunkter.
I dette tilfælde ledte det til en opfattelse af sprogets natur, der ligner det standpunkt jeg lagde frem i kapitel 2.
Hele 'Kundskabens træ' kan ses som en sådan biologisk begrundelse af et epistemologisk standpunkt; det standpunkt om erkendelsens natur som jeg beskrev i starten af kapitlet.
Kapitel 7 Indhold
DISSIPATIVE STRUKTURER
Det har en overgang været almindeligt at betragte livets opståen som en højst usandsynlig hændelse, en unik begivenhed, og dermed det levende som noget højst usædvanligt i universet.
(Fx Monod 71)
Jeg tror livets opståen eller cellens opståen nok er den absolut grundlæggende tilbliven eller emergens (at dukke op); men også at denne tilbliven principielt er af samme karakter som den tilblivelse vi kan observere i vores verden nu.
Jeg vil nu se på om det er berettiget at se tilblivelsen som en højst usandsynlig hændelse; eller om der findes lovmæssige forbindelser mellem de fysiske og de biologiske videnskaber, som kan belyse livets opståen som andet end et mirakel.
Der har i det foregående været et hul mellem vores modeller for det levende og for det døde. De modeller af evolutionen, jeg hidtil har været inde på, har forudsat selvopretholdelse, tilpasningsevne, og reproduktion hos det levende, og kan derfor ikke begrunde disse fundamentale aspekter.
Nyere tiltag indenfor videnskaben giver imidlertid håb om en sådan forbindelse.
(Prigogine 85, s121ff og Bertalanffy 75, s 141f)
7.1 Tidens symmetri og irreversibilitet:
Den klassiske fysik, mekanikken, behandler enkelte partiklers interaktioner, hvor det er karakteristisk at interaktionerne er reversible i tid; de er symmetriske i forhold til tidens retning. Kvantemekanikken behandler enkelte 'partikler' på kvanteniveauet; processerne er symmetriske i tiden, men indeholder også kaos, de er delvist indeterminerede. (Relativitetsteorien er også symmetrisk i tiden.)
Termodynamikken behandler mængder, bestående af enkelte partikler. Går man fra at se på enkelte partikler på kvanteniveauet, til makroskopiske mængder af partikler, sker der et symmetribrud mht. tiden.
Termodynamiske systemer vil bevæge sig i en bestemt retning i tiden, mod en ligevægtssituation, gennem en historisk proces. Det begreb man anvender til at beskrive, hvad der kendetegner systemets ligevægtssituation, kaldes entropi, den irreversible bevægelse mod ligevægt er formuleret i den anden termodynamiske lov: Entropien i et isoleret system kan kun tiltage indtil den termodynamiske ligevægt er nået.
En irreversibel proces er en makroskopisk proces, hvor entropien øges.
Man kan sige at tidens retning er et makroskopisk fænomen; det er de irreversible processer der viser os tidens gang.
Ilaya Prigogine har dog forsøgt at tidens retning fra kvanteniveauet.
Han argumenterer udfra den grundlæggende ubestemthed, det er denne ubestemthed, der ligger til grund for, at irreversibiliteten og dermed tidsopfattelsen findes på det makroskopiske niveau.
Intuitivt kan det forstås således:
Et eksempel på en irreversibel proces er en sten der kastes på en vandoverflade, hvorefter bølger spreder sig ud fra nedslagspunktet; hvis vi kørte en film baglæns, ville en sådan proces afsløre tidens retning for os. På kvanteniveauet sker der det, at en meget stor mængde af partikler bliver korrelerede i et vist omfang, partiklerne bliver forbundet gennem stenens nedslag. Det er disse korrelationer, der danner det mønster, vi ser som bølger, der spreder sig i cirkler og forsvinder. Da hver partikel til en vis grad er ubestemt, er den omvendte proces, med modsat forløb i tiden, ikke tænkbar.
Den ville forudsætte en 'forudkorrelation' af de involverede partikler, partikler langt ude i periferien skulle således være korrelerede i forhold til centret - længe før centret faktisk bliver til.
Dette virker indlysende for den ovennævnte proces, men gælder også for en proces, der kun involverer langt færre partikler. Ubestemtheden umuliggør at man kan tænke sig en verden, der i øvrigt lignede vores, hvor irreversible processer løb i den modsatte retning.
Korrelation mellem en vis mængde partikler kan pga. ubestemtheden kun eksistere i et vist tidsrum, ligesom mønsteret på vandoverfladen langsomt mister sin relation til stennedslaget. Derfor findes irreversibilitet ikke i de enkelte kvantemekaniske processer, men først på det niveau, hvor vi finder korrelationer mellem et vist større antal kvantemekaniske processer.
De korrelationer der ville ses som en omvendt irreversibel proces, ville være af en sådan udstrækning og ikke-forbigående karakter, at det ikke er foreneligt med den grundliggende kvantemekaniske ubestemthed.
(Mere formelt i Prigogine 85, s 303ff)
Den tiltagende entropi kan fortolkes som en bevægelse mod mindre organisation, mod den mest sandsynlige tilstand på kvanteniveauet. Dette ses ofte som en større makroskopisk 'uorden', eller ustruktureret ensartethed.
Hvis et glas koldt og et glas varmt vand blandes, vil molekylernes hastigheder have en fordeling, der består af det varme og det kolde vands hastighedsfordelinger lagt sammen. Molekylerne interagerer og en ny fordeling af hastigheder vil indfinde sig, entropien øges, der indfinder sig et mere sandsynligt udslag af molekylernes samlede bevægelsesenergi. Makroskopisk ses dette som en irreversibel proces, hvor koldt og varmt vand blandes og bliver til lunkent vand.
Form, struktur eller makroskopisk orden kan dog udemærket opstå gennem processer med tiltagende entropi, tænk fx krystaldannelser, dannelse af lermineraler, og 'dråbe'-dannelser lige fra vanddråber til stjernedannelse.
7.2 Orden gennem fluktuation:
Termodynamik har traditionelt beskæftiget sig med isolerede eller lukkede systemer i eller nær ligevægt.
Ligevægt er det eneste referencepunkt for sådanne systemer (som referencepunktet for elektromagnetiske felter og tyngdefelter er partiklens eller jordens centrum).
Ikke-ligevægt har kun været behandlet i forhold til dette referencepunkt.
I de seneste to tiår er man imidlertid blevet mere opmærksom på ikke-ligevægt som en kilde til skabelse af orden og organisation, princippet kaldes orden gennem fluktuation (tilfældigt opståede udsving). Den belgiske fysiker og nobelprisvinder i kemi Ilaya Prigogine har været igangsætter og inspirator for forskningen på dette område.
Et af de fænomener, der har været kendt længe, men som ikke kunne forklares af den gængse termodynamik, er 'Bénards fænomen'. Hvis man tager en væske i termodynamisk ligevægt, og opvarmer den fra neden i en flad skål, vil der opstå en varmestrøm i væsken. Når temperaturforskellene når til et kritisk punkt, dannes der pludseligt et mønster eller en struktur, der ligner biernes 6-kantede celler.
 |
Lignende fænomener er turbulente strømninger og forskellige mønsterdannende og periodiske kemiske reaktioner.
Prigogine kalder sådanne strukturer gennemstrømnings- eller dissipative strukturer (energiforbrugende strukturer); de opstår i åbne systemer, der ikke er i ligevægt. Systemerne inddrager omgivelserne, udveksling af energi og/eller stof er nødvendig for dannelsen af form og struktur; dette kan ses som en basal metabolisme.
Dissipative strukturer har to former for adfærd; tæt på ligevægt destrueres orden, langt fra ligevægt skabes og vedligeholdes orden.
Sådanne strukturer modsiger ikke den anden termodynamiske lov, selvom systemet udviser en negativ entropiændring. Systemet eksporterer blot 'uordenen' eller entropiproduktionen, via sin udveksling med omgivelserne, og sammenlagt vil entropiændringen være nul eller positiv.
I modsætning til ligevægtsstrukturer som krystaller og dråber, kan dissipative strukturer ikke eksistere uden at dannes; at være og at blive til er et og det samme, struktur og funktion er komplementære aspekter.
Heri skjuler sig også termodynamikkens begrænsning overfor fænomenerne; ligesom mekanikken ikke indbefatter irreversibilitet, indbefatter termodynamikken ikke symmetribrud i rummet.
Opretholdelsen af ikke-ligevægt indebærer at der opretholdes en retning i rummet, en makroskopisk gradient for energi og stof. Og netop denne selvopretholdelse, overensstemmelsen mellem en mængde partikler, er et vigtigt aspekt af dissipative strukturer. Opretholdelsen grundes i en form for positiv feedback, selvforstærkning eller selvorganisering i en makroskopisk organisation.
Krystallinske strukturer kan afledes af tiltræknings- og frastødningskræfter mellem enkelte molekyler, og her ud over afspejler de kun kvanteniveauets ubestemthed (fx snefnug).
Til forskel herfra afspejler Bénard-cellerne også den makroskopiske ikke-ligevægt de er oprundet af.
De tre forskellige ovennævnte forklaringsniveauer, kvantemekanik, termodynamik og orden gennem fluktuation, kan ikke reduceres til eller udledes fra hinanden; dette kan forstås udfra de symmetribrud der sker i overgangen mellem niveauerne.
(Jantsch 80, s 22ff)
7.3 Et hierarki af systemaspekter:
Det kan måske give et nyttigt overblik at sætte den dissipative adfærd ind i en sammenhæng med andre systemaspekter.
Systemer karakteriseres efter deres forbindelser med omgivelserne, de kaldes åbne hvis de udveksler energi, stof eller information med deres omgivelser, og hvis de er åbne overfor 'nyheder'.
Udveksling kan kun ske hvis den interne tilstand er ikke-ligevægt.
Den logiske organisation af systemet er processernes karakteristiske sammenhæng i systemet; denne kan være cyklisk eller hypercyklisk (selvkatalyserende).
Organisationen kan være hierarkisk; dvs. bestå af systemer og delsystemer på flere niveauer.
Systemets funktion er den totale karakteristik af systemets processer og forbindelser med omgivelserne. Systemet kaldes autopoietisk når dets funktion primært er selvfornyelse, og systemet først og fremmest refererer til sig selv og ikke noget udenfor.
Strukturen er den faktiske rumlige struktur og kinetik (energitilstand) til et vist tidspunkt. Set over en tidssekvens danner strukturen den totale systemdynamik, systemets 'adfærd'. Systemer afgrænses som tidligere nævnt gennem dynamikken i deres indre organisation.
To helt forskellige typer af systemadfærd kan karakteriseres udfra disse aspekter; bevarende og udviklende adfærd.
(Jantsch 80, s 32ff)
Dette skema viser en mulig hierarkisk ordning af begreber og aspekter omkring systemer:
| Systemaspekter | Bevarende systemadfærd | Udviklende systemadfærd |
| Systemdynamik | Statisk eller bevarende | Dissipativ og nyskabende |
| Struktur | Ligevægtsstruktur | Dissipativ struktur |
| Funktion | Refererer kun til ligevægt | Autopoiese eller selvreference |
| Organisation | Irreversible processer mod ligevægt og tilfældige fluktuationer | Cyklisk forbundne processer og selvkatalyse af fluktuationer |
| Intern tilstand | Nær ligevægt | Ikke-ligevægt |
| Forbindelse til omgivelserne | Isolerede eller åbne | Åbne |
7.4 Fluktuation og stabilitet:
I modsætning til krystaller og andre ligevægtsstrukturer findes der ikke absolut stabilitet i dissipative autopoietiske systemer; systemet kan altid overgå til en ny stabilitetsstruktur gennem fluktuationer.
De betingelser der fører til autopoiesis, åbenhed, ikke-ligevægt og selvforstærkning, ligger også til grund for muligheden for intern forstærkning af fluktuationer; og dermed for videreudvikling.
Autopoiese og udvikling, stabilitet og sammenhængende forandring er komplementære manifestationer af den dissipative selvorganisering.
Fluktuationerne er ikke makroskopiske, men mere eller mindre kaotiske fluktuationer i mekanismer på kvanteniveauet. Nyskabende fluktuationer kan være udslag af perturbationer fra omgivelserne som fx forekomsten af nye reaktionsdeltagere, eller de kan være udslag af interne ændringer i systemet.
Autopoietisk stabilitet repræsenter det specielle tilfælde i et udviklende dynamisk system, hvor fluktuationerne absorberes eller dæmpes i det miljø, de opstår i.
Systemets indre dynamik determinerer emergensen af nye dissipative strukturer, uafhængigt af omgivelserne.
Kun fluktuationer, der når en vis ('kritisk' eller adskillende) udstrækning, fører til at systemet bliver ustabilt og overgår til en kvalitativt ny dynamisk eksistens.
En gennemsnits- eller kvantitativ beskrivelse er ikke gyldig for systemet; fluktuationerne indgår i en stokastisk proces, og den stokastiske skelnen er kvalitativ eller ikke-kontinuert. Meget små fluktuationer 'afgør' systemets fremtidige udvikling; systemet har bestemte muligheder, men den faktiske udvikling er tilfældig, udløst af kaotiske fluktuationer.
Situationen minder om en faseovergang i et fysisk system, hvor mulighederne er bestemte af temperatur, tryk osv., mens den faktiske faseovergang afhænger af en kaotisk fluktuation; dette medfører fænomener, der er problematiske for de videnskabelige modeller, såsom underafkøling og krystallisering.
Sådanne fænomener kan kaldes 'makroskopiske kvantefænomener', et andet aktuelt eksempel herpå er superledning.
Tilfældighed og nødvendighed er komplementære principper i procestænkning.
Fluktuationernes tilfældige effekt afgøres af den nødvendighed, der ligger i koblingen til det øvrige system.
(Jantsch 80, s 42ff)
7.5 Symmetribrud og indeterminisme:
Ved hver overgang til en ny stabilitetsstruktur, vil der være mindst to valgmuligheder for systemet; disse muligheder er symmetriske i rummet, det kan fx være spejlbilleder.
I hver overgang vil der derfor ske et symmetribrud, idet den ene af mulighederne udfoldes, afhængigt af fluktuationernes symmetriforhold.
(Jeg kan ikke lade være med her at tænke på de mange 'valg' af symmetri vi finder i den levende verden.
Et enkelt eksempel er, at der kun findes det ene spejlbillede (L-formen) af aminosyrer (proteinernes byggesten) i levende væsener i vores verden.)
Til forskel fra strukturbevarende mekaniske (fx solsystemet) og termodynamiske systemer, der kan behandles som determinerede på det makroskopiske niveau; støder vi her på den makroskopiske indeterminisme eller ubestemthed, som jeg ofte har agiteret for i det foregående.
Den kvantemekaniske ubestemthed udfoldes i en makroskopisk ubestemthed, i de dissipative strukturers udviklende systemadfærd.
Nogle forskere taler om makroskopisk kvantisering, omkring dannelsen af kvalitativt forskellige dissipative strukturer.
De symmetribrud, der sker i udviklingen af dissipative strukturer, kan ses som makroskopiske kvanteeffekter.
'Orden gennem fluktuation' er en delvist indetermineret og uforudsigelig proces.
Den er derfor også en historisk proces.
Systemer der har gennemgået en udvikling med mange 'valg', mange symmetribrud, fx pga. en stadigt stigende ikke-ligevægt, vil være i én blandt en mængde mulige tilstande.
Hvis systemet nu udsættes for mindsket ikke-ligevægt, vil det gennemgå sin udvikling baglæns, gennem de samme overgange af den samme 'vej'; systemets udvikling er indbefattet eller 'erfaret' i den sidste tilstand. Hver enkelt overgang vil derimod forløbe lidt anderledes i den modsatte retning, den vil ske 'senere' i den faktiske retning, dette fænomen kaldes ofte hysterese (græsk, at blive bagefter). Stabiliteten af en struktur vil altid være større når den er en realitet end når den er en mulighed; - de bestående strukturer er 'overstabile'.
(Jantsch 80, s 47ff)
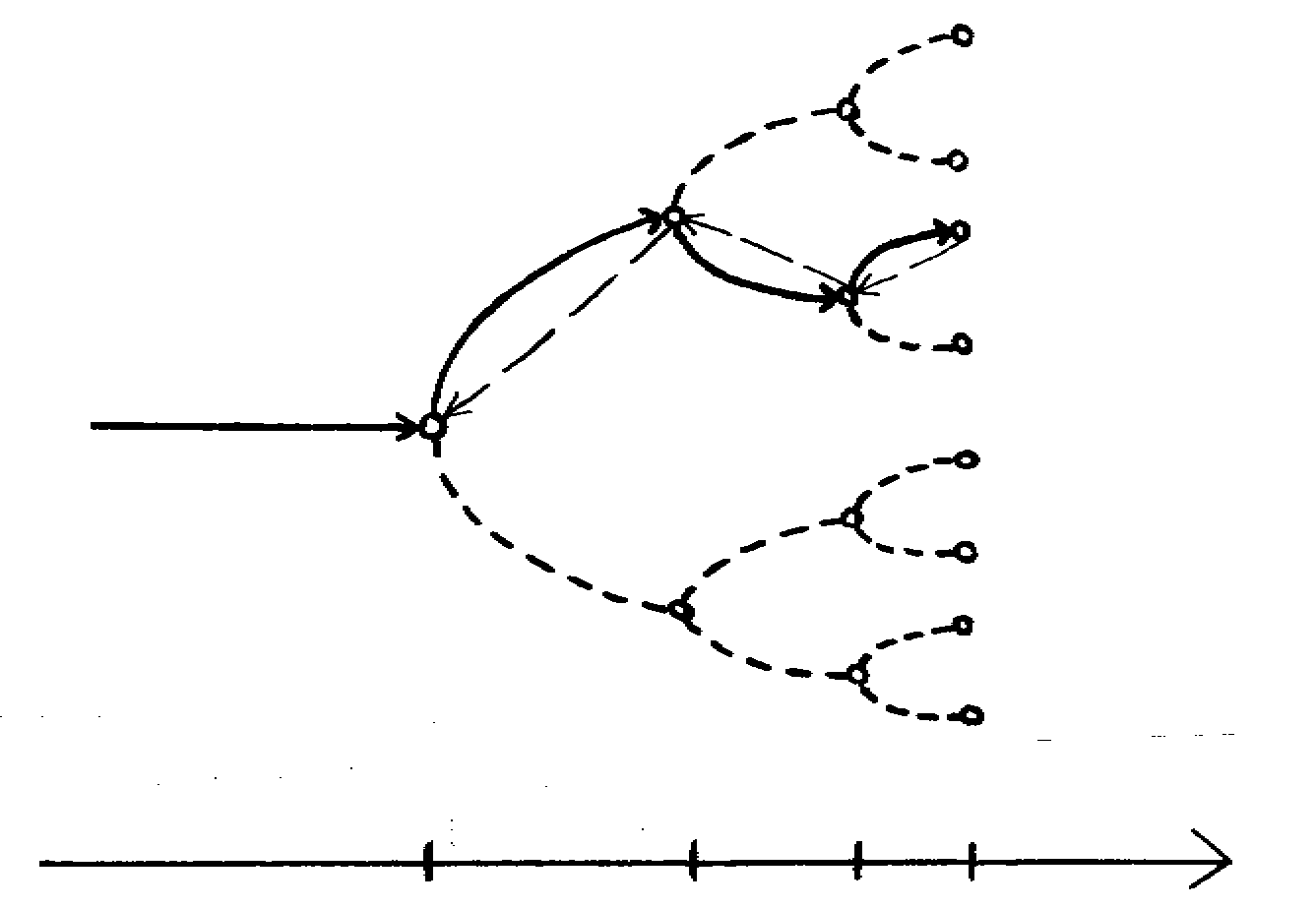
| ||||
| Termodynamisk domæne | 1. Dissipative Struktur (DS) | 2. DS | 3. DS | ... |
Figuren ovenfor viser en faktisk og de mulige dissipative udviklinger, samt hysterese, på en skematisk måde.
(Fra Jantsch 80, s 49)
7.6 Spænding og stabilitet:
Bénards fænomen er et eksempel på, at fluktuationens selvforstærkning til en ny dissipativ struktur følges af en større gennemstrømning af stof og energi (og dermed et højere niveau af interaktion med omgivelserne).
Alle stofstrømme bliver turbulente, hvis strømmens spænding eller potentiale øges udover et vist punkt, og derved øges gennemstrømningen (imt. hvad man umiddelbart tror udfra analogier til turbulens og luftmodstand, men det er fordi man der fokuserer på systemets grænser og ikke på den totale strøm).
Kemiske reaktioner adskiller sig fra de fysiske bl.a ved de helt specifikke reaktionsforhold. Systemets adfærd langt fra ligevægt er specifikt. Instabiliteten opstår ikke kun pga. gradienternes størrelse, hvor selvforstærkningen ligger i en øget strømning; men også gennem nye reaktionsmekanismer, hvor selvforstærkningen ligger i katalytiske sløjfer i reaktionsforløbene.
Stabiliteten af kemiske systemer langt fra ligevægt synes (som for fysiske systemer) at ligge i muligheden for en høj energiudveksling. Mens entropiproduktionen stiger kraftigt i den instabile fase, synes den at nærme sig minimum i en autopoietisk stabil tilstand.
(Prigogine 85, s 190ff og Jantsch 80, s 43 og 50)
En anden indgang til fænomener, der minder om instabile overgange mellem dissipative strukturer, er den matematiske katastrofeteori der er udviklet af René Thom.
Systemernes adfærd beskrives her ved hjælp af geometriske modeller, der fokuserer på katastroferne, på overgangene mellem stabile tilstande. Aspekter som selvopretholdelse indgår ikke i modellerne, men de beskriver opståen af ny form i lyset af delvis selvdeterminering. Sagt på en anden måde belyser katastrofeteorien ikke-kontinuerte effekter af kontinuerte ændringer.
Et simpelt mekanisk eksperiment, der kan udføres af enhver, kan give et billede af katastrofernes natur, og det er faktisk ganske fascinerende.
Som figuren viser, består eksperimentet i en vekselvirkning mellem to kræfter, fjederkræfterne i de to elastikker. Papskiven skal kunne dreje let om centret.
 |
7.7 Information; nyhed og bekræftelse:
Information har været tolket som negativ entropi (negentropi); altså som rumlige eller tidslige relationer, organisation eller orden.
Men som Bateson definerer information (en forskel der gør en forskel) er det karakteristiske ikke orden men effekt; information er noget der har en indvirkning.
Information i denne forstand kaldes også pragmatisk (effektiv, virkende eller handlende) information. Pragmatisk information har ingen selvstændig eksistens, den er 'kun' et aspekt af en informationsproces.
Til forskel fra negentropi er pragmatisk information sammensat af to komplementære aspekter; kaos og orden eller nyhed og bekræftelse.
Ren nyhed, det unikke, står for kaos; det indeholder intet der kan virke, for virkning forudsætter genkendelse eller overensstemmelse.
Ren bekræftelse bringer intet nyt, kun stagnation; ren bekræftelse har ingen effekt, fuld overensstemmelse bringer ingen forandringer.
Informationens virkning findes derfor i en balance mellem nyheder og bekræftelser.
Nedenstående figur viser denne balance (meget) skematisk:
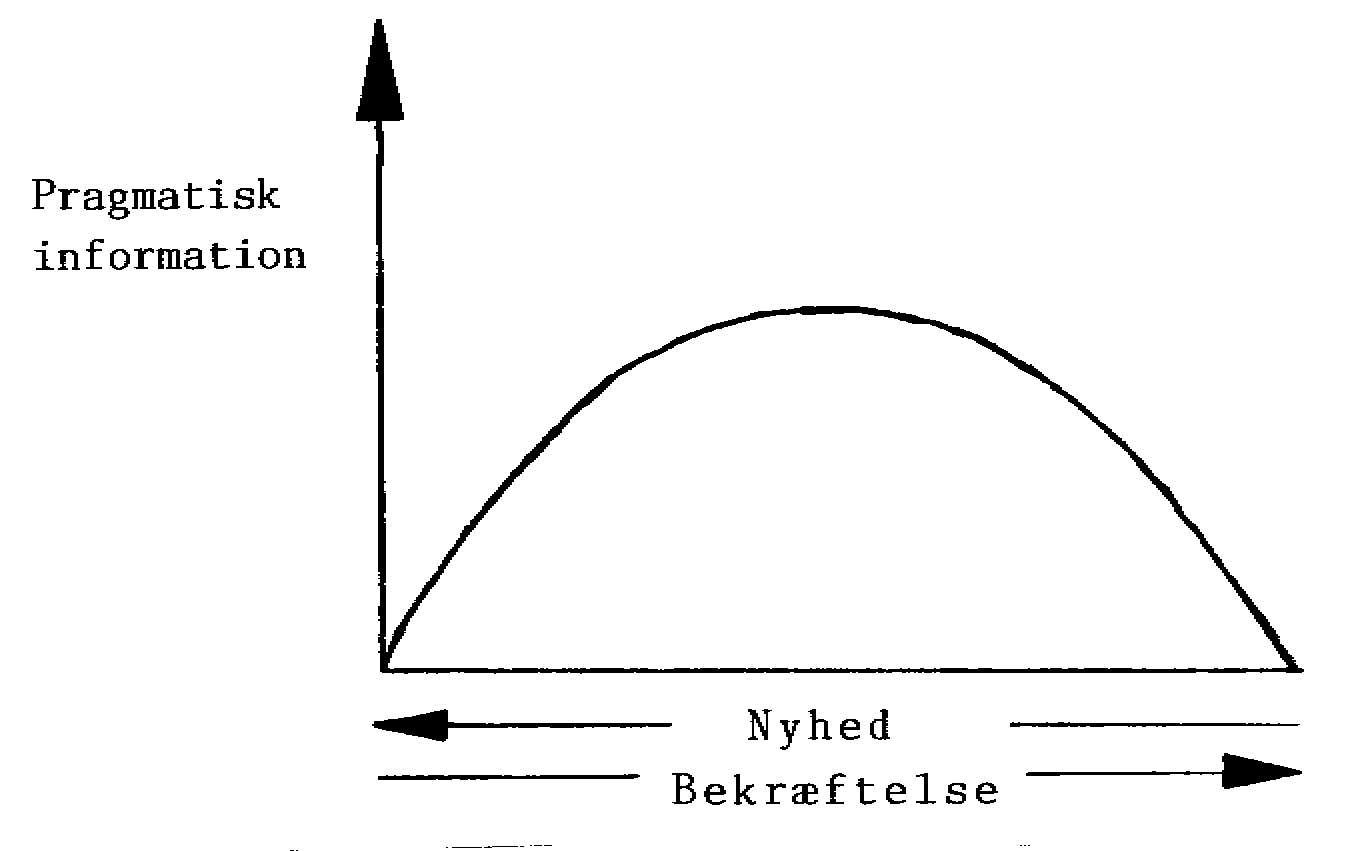
(Efter Jantsch 80, s 52
Den konventionelle matematisk formulerede informationsteori, grundlagt af Shannon og Weaver, betragter ligevægts- og bekræftelsesaspekter; analogt med den konventionelle termodynamik. Teorien er derfor kun gældende for den mest bekræftende information, længst til højre på figuren.
Vi kan nu etablere en forbindelse mellem vores model af pragmatisk (virkende) information og de ordnende principper i ligevægts- og ikke-ligevægtsstrukturer.
Ren bekræftelse svarer som nævnt til stagnation og dermed termodynamisk ligevægt, i ligevægt absorberes og forsvinder enhver perturbation, den pragmatiske information er nul.
På den anden side svarer ren nyhed til kaos, dette kan fortolkes som instabilitetstærsklen i en dissipativ overgang.
De stokastiske processer (der sammenligner, dvs. 'vælger' det bekræftende) refererer ikke længere til den gamle struktur, og den nye er endnu ikke opstået. Den pragmatiske information er også her nul, ikke fordi perturbationerne forsvinder, men fordi det efterfølgende resultat er ubestemt.
Den autopoietiske stabile ikke-ligevægt, de dissipative strukturer må findes imellem ligevægt og ustabilitet, i en balance mellem nyhed og bekræftelse omkring den pragmatiske informations maksimum.
(Jantsch 80, s 50ff)
I den nedenstående figur er der, foruden ligevægtsstrukturer, skitseret en dissipativ strukturs udvikling over instabilitetstærsklen til en ny autopoietisk stabil tilstand.
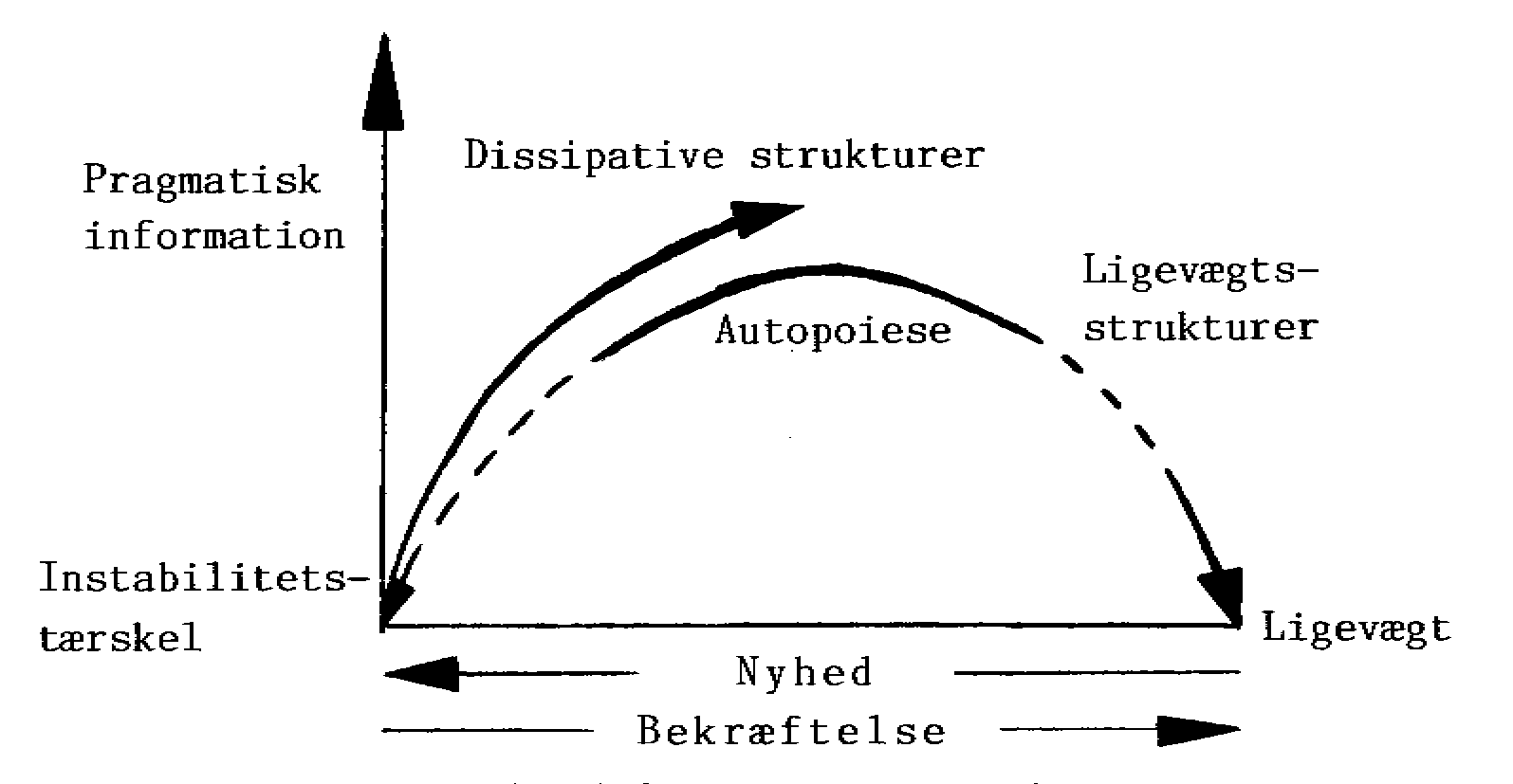 |
Jeg tror, at teorien om dissipative strukturer må indgå i en fremtidig dynamisk teori om historiske systemer; systemer hvis udvikling afhænger af hvert af dets undersystemers historie. En generel model for systemer som helheder, der kan hjælpe os at forstå fluktuationer og stabilitet i biologiske og sociale systemer, og aspekter af virkeligheden som organisation, hierarki, epigenese, strukturelle koblinger mv.
Det er ikke sikkert at den ovenfor skitserede forbindelse mellem termodynamik og informationsteori er den rette; men jeg tror, at der findes en sådan forbindelse og, at den må befinde sig centralt i en generel dynamisk/historisk systemteori.
(Se også Bertalanffy 75, bl.a s 149ff)
Kapitel 8 Indhold
DEN HISTORISKE EVOLUTION
8.1 Darwin og historiske processer:
Jeg har tidligere været inde på de problemer der er forbundet med ekstrapolationer, og specielt ekstrapolationer i historiske processer.
Jeg kan ikke uden videre acceptere neodarwinismens opfattelse af makroevolutionen som en simpel ekstrapolation af de mikroevolutionære mekanismer. Det ville i hvert fald være reduktionistisk ikke at undersøge, om der findes andre regelmæssigheder i den historiske evolution vores klode har gennemlevet.
Darwin formåede at overbevise mange af sine samtidige om, at jorden havde gennemgået en organisk evolution, selvom hans forslag til en drivende mekanisme ikke nød den samme tiltro.
Darwins argumenter er stadig overbevisende, og en god grund hertil kan findes i den metode Darwin benyttede, da han stod overfor problemerne med at behandle historiske processer på en videnskabelig måde.
Det vigtigste problem er (som omtalt i afsnit 3.5), at vi kun kan iagttage historien i vores egen tidshorisont, lige nu.
De processer, der har frembragt de tilstande vi ser nu, kan vi kun iagttage mere eller mindre indirekte.
Darwins historiske metode kan udmøntes i tre principper, der hver for sig kommer frem i tre mindre kendte bøger som Darwin skrev.
Bøgerne behandlede tilsyneladende små og begrænsede emner i naturhistorien; regnormes betydning i dannelsen af muld, koralrevs struktur og udbredelse, og insekters bestøvning af orkideer.
Regnormene belyser det princip, at små og ubetydelige men målbare processer kan være årsag til overordentlig betydelige effekter.
Dette princip bygger som de andre på at man indser eller overvejer den dynamiske natur af tilsyneladende statiske tilstande, som her muldlaget.
Tegningen nedenfor viser Darwins teori om koraløers og atollers dannelse gennem gradvis sænkning af havbunden og samtidig vækst af korallerne:
(Her fra Bonde 85, s 245)
Koralrevenes vækst og den synkende (eller rettere skrumpende pga. afkøling) havbund kan ikke måles i vores korte tidshorisont. Men hvis sådanne processer er startet til forskellige tider, eller løber med forskellige hastigheder; vil der findes eksempler der befinder sig på forskellige stadier i processens forløb.
Vi kan da forsøge at konstruere en teori, der kan forklare eller udlægge disse eksempler som stadier i en enkelt historisk proces. Et velkendt eksempel på anvendelsen af denne helt generelle metode er undersøgelsen af stjernernes livsforløb.
Orkideerne giver eksempler på tilsyneladende statiske objekter, der fremviser deres historiske karakter i de mærkværdigheder, der netop synes mærkværdige fordi de ikke lader sig forklare udfra det nuværende; mærkværdighederne må begrundes i tidligere tilstande og kontekster, i orkideernes historie.
Netop dette princip giver nogle af de vægtigste indicier for en evolution, hvori der indgår en ikke-almægtig naturlig selektion.
(Her fra Gould 83b, s 120ff)
Francois Jacob skriver i sin bog 'Mulighedernes spil':
"Komplekse objekter, hvad enten de er levende eller ikke, er altid produkter af evolutionsprocesser, hvori to faktorer medvirker: dels de tvingende lovmæssigheder, der på hvert niveau bestemmer spillets regler og markerer det muliges grænser; dels de omstændigheder, der styrer begivenhedernes faktiske retning, og som omsætter systemernes interaktioner i virkelige forhold. Kombinationen af lovmæssigheder og historie genfindes for hvert niveau, men i forskellige blandingsforhold."
(Jacob 85, s 67)
Eller som jeg tidligere har formuleret det, tilfældighed og kaos er tilstede på alle niveauer i vores verden.
Den biologiske verden er ikke perfekt eller fuldkommen, som den ville være hvis en altformående forudseende ingeniør havde skabt den.
Levende væsener ligner snarere produkter fra en fingernem hobbyfusker der arbejder i nuet og med hvad der tilfældigt er for hånden, uden opskrift eller konstruktionsplan.
En sådan hobbyfusker anvender ofte det givne på nye og ikke forudsete måder. Jacob kalder denne proces for bricolage.
Darwin så, at fx vigtige strukturer i orkideernes befrugtning lignede resultatet af en sådan bricolage, udarbejdet udfra på forhånd eksisterende organer der oprindeligt havde en bestemt opgave, men som nu havde andre funktioner.
I Arternes Oprindelse myldrer det med eksempler af denne art.
Bricolage er dog stadig en menneskelig adfærd, og indeholder derfor betydninger, der ikke gælder for evolutionsprocesserne. Hobbymanden vil ofte danner sig billeder af resultatet af bricolagen; han arbejder delvist finalt eller målrettet. Naturlig udvælgelse eller naturlig drift kan kun ske udfra de koblinger der er tilstede, ikke udfra de der kunne komme, hvis... osv.
Evolutionen er mulige omdannelser af det eksisterende, og kaos og begrænsninger i kaos spiller begge en rolle.
Kaos er skabelsen af det nye; begrænsningerne er mange og beskrives som fysiske, kemiske, biologiske og evolutionære lovmæssigheder.
Stephen Jay Gould skriver herom:
"Organismer arver en kropsform og en speciel type epigenese; disse begrænser fremtidige ændringer og tilpasninger.
I mange tilfælde reflekterer udviklingsveje de arvelige mønstre mere end fordringer fra omgivelserne. Det arvede begrænser, men tilbyder også muligheder. ... Evolutionens fleksibilitet udgår måske primært fra sideeffekter uden tilpasningsværdi, der gør det muligt for organismer at gå i nye og uforudsigelige retninger."
"At forveksle nuværende nytte med grunden til den historiske oprindelse er en logisk fælde, der har plaget evolutionær tænkning fra starten."
(Gould 83b, s 156 og 170)
Begrebet tilpasning bør bruges med forsigtighed; som jeg var inde på tidligt i opgaven, vil en ukritisk brug føre til ringslutninger og begrebet mister sin nyttighed.
Maturanas 'strukturelle kobling' gør bedre opmærksom på den dynamiske gensidige relation, der er grundlaget for de tilpasninger eller strukturelle overensstemmelser vi kan undersøge; og indeholder ikke den målrettethed der kan ligge i 'tilpasning'.
Når jeg bruger begrebet tilpasning, og det er ikke let at kassere et så anvendt begreb, tænker jeg derfor på en proces der minder om strukturel kobling, og tilstande der ligner strukturel overensstemmelse.
8.3 Tilfældighedens rolle:
Jeg vil her kort ridse tilfældighedens rolle op på de lavere niveauer i evolutionen, populationens forandringer og artsdannelsen, som baggrund for det større perspektiv i næste afsnit. (Jeg mener her tilfældighed i forhold til den naturlige udvælgelse på organismerne.)
Den genotypiske variation i populationen, populationens fleksibilitet, er betinget af individernes fleksibilitet (se afsnit 5.5). Populationens genetiske sammensætning er derfor, dels pga. individernes fleksibilitet og dels pga. genomets indhold af 'overflødigt' DNA, kun til en vis grad 'synlig' eller til stede for den naturlige udvælgelse.
At det faktisk forholder sig sådan, antydes af undersøgelser, der viser en meget stor genetisk variation i naturlige populationer. Hos diploide organismer vil en stor bestanddel unyttig eller skadelig variation også kunne 'skjules', fordi organismen har et dobbelt sæt genetisk materiale. (Dette er nok den vigtigste årsag til at indavl, der fører til at de to sæt bliver mere ens, ofte er skadelig.)
Udbredelsen af variation, der ikke styres af strukturelle koblinger til omgivelserne, er delvist tilfældig (genetisk drift); men påvirkes også af interne koblinger, som fysisk nærhed til mere eller mindre stabile dele af kromosomerne, og en indre overensstemmelse med organisation og epigenese (fx i meiosen, se afsnit 5.3).
Afgrænsningen af arten bygger i princippet på reproduktiv sammenhæng indenfor arten, og reproduktiv isolation mellem arter. Udfra denne opdeling bliver det centrale spørgsmål i artsdannelsen: Hvordan og hvorfor opstår den reproduktive isolation?
Traditionelt har man troet på dannelsen af geografiske barrierer og en efterfølgende opsplittende udvikling styret af naturlig udvælgelse, hvorved reproduktiv isolation kan opstå som et biprodukt. Nyere teorier foreslår at den reproduktive isolation kan opstå spontant, som et resultat af historiske tilfælde i genomet, uden betydning for tilpasningen. Og endvidere at disse tilfældigheder skaber de grænser, der giver naturlig udvælgelse lejlighed til at fungere som en opsplittende mekanisme i populationen. Altså et udgangspunkt, der er tilfældigt i forhold til de ydre omstændigheder for artsdannelsen.
Jeg vil i næste kapitel behandle artsdannelsen mere indgående.
8.4 Tilfældighedens rolle i et større perspektiv:
De fossile data fortæller os, at forskellige typer af organismer har været udbredte i forskellige epoker af livets historie.
Den almindelige opfattelse er at sådanne makroevolutionære ændringer afspejler den naturlige udvælgelse i stor skala; krybdyrene blev afløst af deres bedre tilpassede efterkommere pattedyrene, trilobitterne uddøde og må derfor have været dårligere tilpassede end de efterfølgende krebsdyr.
Men disse skift har ikke været gradvise; jordens historie er blevet opdelt i epoker, Kul, Perm, Kridt, osv., og disse epokers grænser er ofte sammenfaldende med markante perioder af masseuddøen, der skiller sig ud fra de mellemliggende perioder (eller – epokerne er inddelt efter disse skift).
Nyere undersøgelser har nærmere analyseret uddøen af familier (en systematisk kategori over slægten) i de sidste 600 millioner år. (Pga. de meget fåtallige fossiler, kan analysen ikke foregå på artsniveau.)
Den 'normale' uddøen er 2-4 familier per mio. år, mens den i masseuddøensperioder er op til 20 familier per mio. år.
Den største masseuddøen skete for omkring 225 mio. år siden. Halvdelen af de marine organismers familier forsvandt i denne periode, heriblandt de sidste trilobitter. Forskellige forskere anslår at det er ensbetydende med, at mellem 80 og 95 procent af jordens arter uddøde i løbet af nogle få millioner år. I slutningen af kridt, da også dinosaurerne forsvandt efter flere hundrede mio. års dominans, anslås det at 75 procent af alle arter uddøde.
(Man har fundet spor af en flercellet fauna, der levede før den velkendte opblomstring af flercellede organismer for ca.600 millioner år siden hvor stamformerne til de nulevende flercellede opstod. Denne fauna, der havde en fladeformet opbygning der er helt forskellig fra de nuværende flercellede dyrs rørformede opbygning, uddøde fuldstændigt og det er måske den mest drastiske masseuddøen i livets historie.)
(Bl.a Gould 85, s 236ff og Gould 78, s 136)
Der har været foreslået mange forskellige forklaringer på hvorfor så mange arter i vidt forskellige miljøer uddøde i bestemte perioder, bl.a kontinenternes indbyrdes placering, ændringer i havniveauet, og senest, at jorden blev ramt af en asteroide eller enorme vulkanudbrud.
Det er i denne forbindelse lige meget hvad der udløste de globale kollapser; det interessante er selve forekomsten af masseuddøen. For det fortæller os, at de større mønstre i evolutionens forløb ikke kan forklares udelukkende ud fra de strukturelle koblinger vi kan undersøge i dag.
Et billede fra økologien kan vise hvad jeg mener.
I en sø eksisterer et væld af koblinger og interaktioner, som vi kan undersøge og forbinde med de forskellige arter og populationers udbredelse.
Men hvis søen bliver voldsomt eutrofieret (næringsstofforurenet) i løbet af kort tid, vil de undersøgte koblinger ikke være relevante for den nye situation; aborren er tilpasset til en 'normal' situation, ikke til menneskeskabt forurening.
Tilsvarende må uddøen i en masseuddøensperiode (og den form for naturlig udvælgelse der indgår) være delvist tilfældig i forhold til den naturlige udvælgelse af individer, der er foregået i den forudgående 'normale' periode, og de interaktioner vi kalder konkurrence, predation, symbiose osv.
Eller sagt på en anden måde, netværket af strukturelle koblinger fungerer i en vis kontekst af miljøvariable; sker der en ikke-tolerabel ændring i en eller flere miljøvariable (fx temperatur, 'plads' eller lysstyrke) vil de omvæltninger, der sker i netværket (den masseuddøen vi kan registrere), ikke kunne forklares udfra de forestillinger vi kan danne os om de 'normale' strukturelle koblinger.
Det er kun få procent af kridttidens arter, der har nulevende efterkommere, men det betyder ikke, at hovedparten af kridttidens arter var dårligt tilpassede til deres miljø!
Ligeledes kan vi ikke forklare den varierende udstråling eller forgrening af de tilbageværende former, som vi kan konstatere efter en periode med masseuddøen, blot som et produkt af individernes og populationernes tilpasning til deres omgivelser. De forskellige former må antages at være forskellige også med hensyn til deres potentiel for artsdannelse.
Helt generelt kan vi altså sige, at der i makroevolutionære ændringer i og imellem klader (af græsk klados, grenafsnit) indgår en variation i frekvens eller hastighed af artsdannelse og uddøen, der ikke er bestemt af arternes tilpasning til deres omgivelser.
Det indgår i den hierarkiske opfattelse, at reglerne eller lovmæssighederne på forskellige niveauer er af forskellig natur.
Vi kan derfor ikke betragte den historiske evolution som en simpel ekstrapolering af individernes naturlige udvælgelse, men må være forberedt på, at de strukturelle koblinger, der spiller ind på evolutionens forskellige niveauer, ikke er de samme.
Delfinens eminente tilpasning til dens nuværende omgivelser kan ikke sige os noget om, hvorvidt delfinens efterkommere vil 'få succes' i et større evolutionært perspektiv.
8.5 Kladistisk systematisering af mønstre:
Resultaterne af en historisk proces kan systematiseres ved hjælp af en kladistisk metode.
Metoden går egentlig ud på at finde specielle ligheder (synapomorphier), dvs. ligheder der er specielle for de senest opsplittede eller tættest beslægtede enheder, kaldet søstergrupper. Og der udfra konstruere et billede af mulige afstamningsslægtskaber mellem enhederne (dette ser jeg på i næste afsnit).
I bestræbelserne på at gøre kladistikken så strengt videnskabelig som muligt, foretages sammenligningerne imidlertid helt uden henvisninger til den historiske proces.
Man sammenligner et meget stort antal 'karakterer' (ved hjælp af datamaskiner) uden at skelne mellem nye, specielle og ældre, almene ligheder eller ligheder uafhængigt af slægtskab; udfra den antagelse at specielle ligheder er de talrigste, og derfor afgørende når mange karakterer sammenlignes. De undersøgte karakterer kan fx være molekylære data, udfra analyser af proteiner og DNA.
Molekylære data har den fordel frem for makroskopiske træk, der kan ligne ubeslægtede træk pga. konvergent (mod det samme) tilpasning, at en stor del af ændringerne ikke har betydning for tilpasningen. Lowenstein angiver, at 95% af ændringerne i proteiner, og 99% af ændringerne i DNA er tilpasningsneutrale. Det er også dette, der ligger bag det såkaldte molekylære ur; antallet af molekylære forskelle mellem organismer korrelerer med afstanden til tidspunktet for deres opsplitning.
(Lowenstein 85, s 544)
Kladisternes udgangspunkt er derfor først og fremmest de nulevende dyr, idet et fossil altid kun er en (meget) begrænset afbildning af et levende væsen, og derfor ikke kan indgå på lige fod i kladistiske sammenligninger af et stort antal karakterer.
Denne 'data'-kladistik giver en systematisk sammenhæng mellem nulevende former. Den danner derfor en ramme, hvorimod man kan afprøve andre undersøgelser af evolutionens mekanismer og det faktiske fylogenetiske forløb.
Mere generelt kan man se kladistik som en systematisering af det mønster af molekylære, anatomiske og adfærdsmæssige ligheder som er fremkommet gennem evolutionen; mønsteret er et 'tværsnit' af den historiske proces.
På lignende vis danner den rumlige fordeling af populationer, arter og højere klader et mønster, en anden type tværsnit af evolutionen (teorien om kontinentaldrift kan være med til at forklare disse biogeografiske mønstre).
Strukturelle koblinger og økologiske sammenhænge giver endnu et tværsnit af den historiske proces.
Systematisering af disse mønstre danner en nødvendig baggrund for en analyse af evolutionen som proces, og en forståelse af lovmæssigheder og tilfældighed i fylogenien.
(Bl.a Cracraft 83, s 273ff)
Darwins principper gjaldt analysen af sådanne mønstre; udfra antagelser om sammenhænge mellem forskellig typer koralrev, mellem orkideer og andre blomster, og mellem regnormes liv og muld kunne han danne hypoteser om den historiske proces.
De forskelle og ligheder vi systematiserer i vores 'tværsnit' antages at være et billede af de forandringer, der tilsammen udgør den historiske proces.
Men forskelle og ligheder siger ikke noget om den tidslige retning og rækkefølge forandringerne er sket i.
Darwins forslag til en analyse heraf var: at måle processen direkte; at fastlægge tidens retning via de irreversible processer der indgår; og at se på mærkværdige strukturer, der kun kan forklares i en bestemt tidslig sammenhæng.
8.6 Kladistisk analyse og fylogenier:
I den kladistiske analyse er formuleret bestemte metoder til at udskille specielle ligheder, fastlægge tidens retning, og derved rekonstruere eller teste fylogenier; hvilket ikke er muligt med den systematiserende kladistik som jeg omtalte ovenfor.
'Outgroup'-kriteriet kaldes således, fordi karakterer eller modifikationer der deles med individer uden for (men beslægtede med) den gruppe man undersøger, må regnes for tidligere end gruppens opsplitning; og altså er en generel eller oprindelig lighed.
I virkeligheden vil der oftest være modstrid mellem analysen af forskellige karakterer; nogle modifikationer kan let dannes og tilbagedannes, mens andre modifikationer er ret irreversible.
Oftest vil man foreløbigt antage den simpleste model blandt de mulige, den model der foreslår det mindst mulige antal tilbagedannelser efter et sparsomhedsprincip, fordi vores viden om forskellige forandringers nyttighed i en rekonstruktion af fylogenien ikke er tilstrækkelig.
Nogle modifikationer, fx de mere komplekse der er uafhængige af strukturelle koblinger, er dog af større værdi end andre som indicier for en fælles afstamning, og imod en analog udvikling.
Det ontogenetiske kriterium bygger på, at forandringerne i ontogenesen i et vist omfang korrelerer med forandringer i fylogenesen.
Det epigenetiske forløb er karakteriseret ved at være en udvikling der bygger på det forudgående. Dette indebærer, at en genotypisk forandring vil have større konsekvenser jo før i epigenesen den indvirker; og på ethvert fylogenetisk tidspunkt vil de forandringer, der indvirker sent i epigenesen, have større chance for at kunne indpasses i en fungerende organisme.
Fylogenetiske forandringer må ske gennem ændringer i individernes epigenese. Det er således resultatet af strukturelle koblinger i epigenesen, der viser sig som den generelle regel at ontogenetiske ændringer vil ske fra fylogenetisk ældre til fylogenetisk yngre karakterer.
Brugbarheden af denne metode er begrænset til de karakterer der kan identificeres i ontogenesen, og dermed til den nyere del af de flercellede organismernes fylogenese. Vanskeligheder findes fx også med insekternes metamorfose, der er en drastisk rumlig omfordeling i ontogenesen.
Desuden vil genotypiske forandringer, der bevirker at forskellige dele af den epigenetiske proces forskydes i tid i forhold til hinanden, udgøre et problem for det ontogenetiske kriterium.
Et velkendt eksempel er, at evnen til at forplante sig kan optræde på et tidligere ontogenetisk stadie, således at de senere stadier måske slet ikke kommer til udtryk.
Dette kendes fra padder, som axolotlen, der forplanter sig i 'haletudse'-stadiet, og fænomenet udgør et væsentligt indlæg i argumentationen omkring menneskets udviklingshistorie.
(Gould 77b, s 319ff)
Palæontologiske data (fossiler) kan ikke i sig selv danne et billede af evolutionens forløb, dertil er deres afbildning af de levende organismer for begrænset og de fossile data for mangelfulde; men de kan give konkrete bidrag til et billede af evolutionen, hvis de tolkes i lyset af de øvrige kladistiske metoder. Desuden udgør palæontologien vores eneste mulighed for at kalibrere vores kladistiske analyse af tidsforløb med det faktiske tidsforløb i den historiske evolution.
(Bl.a fra Bonde 87, s 144ff og Cracraft 83, s 273ff)
8.7 Slags og slægtskab:
Der viser sig et helt generelt klassifikationsproblem når vi undersøger historiske processer.
Klassifikation efter fylogenetisk slægtskab stemmer sjældent overens med de klassifikationer vi lægger ned over enhederne udfra kriterier om 'slags', kriterier der bygger på form og funktion.
(Gould 83b, s 364ff)
Vi stødte på problemet allerede i definitionen af artsbegrebet, hvor konflikten opstår mellem reproduktionskriteriet og det økologiske nichekriterium. Det er interessant at man har forsøgt at definere arten udelukkende udfra kriterier om slægtskab, mens begreber som tilpasning og overensstemmelse hører til opdelingen i slags.
I den mere overordnede systematiske klassifikation er konflikten så at sige indbygget; idet den grundlæggende ramme for fylogenetiske forslag, den kladistiske systematik, slet ikke refererer til fylogeni.
Denne tilsyneladende konflikt er dog faktisk styrken i kladistik, idet man derved har opnået et uafhængigt grundlag for afprøvning af de fylogenetiske hypoteser. De andre 'tværsnit' af udviklingsprocessen, fx biogeografiske og økologiske data er andre mulige rammer for afprøvning af slægtskabshypoteser.
Vi har dermed mulighed for at vurdere vores umiddelbare inddeling af de levende væsener i forskellige slags, fx menneske og menneskeaber, i et udviklingshistorisk perspektiv.
Vores klassifikationsproblem tjener også til at vise os, at den underliggende historiske proces er forskelligartet; fordelingen af stasis og forandring er 'ujævn', og forandringerne er af forskellig type eller sker i forskelligt tempo.
Det interessante ligger i at prøve at forstå denne forskelligartethed, at belyse de lovmæssigheder der findes og de historiske tilfælde der finder sted.
Ovennævnte konflikt udgør endnu et eksempel på, at enhver inddeling er vilkårlig og at der findes mange muligheder for opdeling.
Lidt mere systematisk kan man stille vore muligheder for opdeling af de levende væsener på denne måde:
Man kan fx betragte to forskellige inddelinger i slags:
Kladistisk systematik - udfra mikroskopiske ligheder i form.
Sædvanlige 'slags' - (makroskopiske) ligheder i form og funktion (her er mange mulige underopdelinger).
Der er også flere niveauer i vores inddelinger i slægt, fx:
Sociale koblinger - reproduktion og populationer.
Kladistisk analyse - specielle slægtskabsligheder.
Fylogenier (stamtræer) - forslag til faktiske rækkefølger og forgreninger, palæontologiske data.
Evolutionshistorie - større sammenhænge mellem klader.
Kapitel 9 Indhold
EVOLUTIONENS MEKANISMER
De overvejelser jeg fremsætter i dette kapitel må af nødvendighed være spredte og forsøgsvise. Evolutionens mekanismer er endnu kun dårligt forståede, og for nogle dele af det levende endnu mindre end det.
Jeg vil i gennemgangen benytte de synsvinkler jeg er kommet frem til i opgaven, og ikke knytte an til nogen bestemt evolutionsteoretisk opfattelse.
9.1 Kontekst og evolution:
Når man snakker om hændelser og udvikling, er det almindeligt at sige, at hændelsen sker 'i' en vis sammenhæng eller kontekst.
Denne måde at snakke på giver det indtryk at konteksten er uafhængig af og bestemmende for hændelsen, - og skjuler dermed at konteksten netop består af sådanne forbindelser som hændelsen repræsenterer.
Hestens udvikling er en af de virkelig velunderbyggede fylogenier. Det ville være forkert at se denne udvikling som en række forandringer i dyrets tilpasning til livet på græssletterne.
Græssletterne selv udvikledes i samme takt til græsæderne; hestens og græssletternes udvikling sker i og er bestanddele af en fælles kontekst.
'Overlevelse' betyder, at visse beskrivende udsagn om et levende system fortsætter med at være sande i en tidsperiode; 'evolution' refererer til forandringer i sandheden af visse beskrivende udsagn om et levende system.
Problemet er at definere, hvilke udsagn om hvilke systemer der forbliver sande eller undergår forandringer.
I hestens tilfælde overlever forholdet mellem dyr og miljø i et længere perspektiv, mens hest og græs undergår forandringer og tilpasser sig fra øjeblik til øjeblik.
Dette princip er blevet kaldt 'Den røde dronnings hypotese' efter en af Lewis Carrolls figurer, der siger til Alice:
"Se her, det kræver at du løber alt hvad du kan, for at blive på samme sted."
Tilpasninger er ikke hele historien.
Hvis forholdet mellem dyr og miljø skal overleve, må ændringerne hos den ene part modsvares af ændringer hos den anden. Nogle ændringer i en sådan systemproces fremtræder som en slags paradokser (hændelser der virker paradoksale udfra reglerne på det niveau hvor de sker), fordi reglerne for ændringerne er forskellige fra reglerne for overlevelse og udvikling af det økologiske system; de økologiske sammenhænge eksisterer i en længere tidshorisont end de deltagende arter.
Hændelser der fremtræder som paradokser kan fx være artsdannelser og uddøen. På andre niveauer i evolutionen kan man støde på lignende paradoksale hændelser; fx altruisme, individets aldring og organismens død gennem grænseoverskridende adaptationer; eller de forskellige økologiske koblinger der virker paradoksale fra organismernes niveau, hvis man ikke forstår reglerne for de overlejrende økologiske sammenhænge.
Et vist systems kontekst, den sammenhæng det eksisterer i og som en del af, må tilhøre en større tidshorisont (og/eller rumlig horisont) end systemet; dette er en forudsætning for at vi kan betragte det ene som kontekst for det andet. Linedanseren danser på linen, dette synspunkt indeholder at balancen nødvendigvis må befinde sig i en længere tidshorisont end dansen; eller man kan sige at reguleringen af balancen overlejrer reguleringen af dansen.
(Bl.a Bateson 85, s 338f)
9.2 Forandring og konstans:
De problemer, jeg tidligere har været inde på omkring slags og slægtskab og mere eller mindre springvis evolution, viser sig specielt tydeligt i diskussioner om artsdannelse.
Udover de indvirkende evolutionære mekanismer afhænger artsdannelsen som proces af vores opfattelse af arten som et led i denne proces.
Det kan ofte være frugtbart at fastlægge et begreb som arten af hensyn til den videnskabelige produktivitet.
På den anden side er det nødvendigt for den videnskabelige forståelse regelmæssigt at tage sådanne begreber op til overvejelse.
Jeg har tidligere været inde på de problemer der findes i at definere artsbegrebet uden henvisning til evolutionen. Selv hvis vi kun ser på organismer med seksuel formering, er der problemer med at sætte skel mellem 'rigtige' arter, underarter og varieteter.
Dette kan skyldes at vi betragter et tværsnit af processen, og derfor både ser de egentlige enheder og overgangssituationer, som når en fiksering af en bakteriekoloni viser stadier i delingen. Men det kan også skyldes, at enhederne mere findes i vores opfattelse end i selve processen.
Ser vi på artsbegrebet i evolutionens tidshorisont, har vi dels de kladistiske holdepunkter, forgreningerne; de nulevende arters opsplitning kan fastsættes med ret stor sikkerhed udfra den molekylære kladistik og en palæontologisk kalibrering. På den anden side har vi forandringerne i slags, i form og funktion; det umiddelbare grundlag for vores opdeling af verden.
Er arten en stabil enhed?
Som Bateson sagde, må vi forsøge at definere hvilke udsagn om hvilke systemer der forandres og hvilke der vedbliver at være sande. Et sådant spørgsmål kan derfor ikke besvares med ja eller nej. Meget få aspekter af det levende har været stabile igennem hele evolutionens historie.
Stabilitet er et begreb der forudsætter en tidshorisont.
Og det vil kun være nogle udsagn om artens organismer, der besidder denne stabilitet, og vedbliver at være sande.
I denne sammenhæng, er det nyttigt at bruge en opdeling af organismernes funktion i et socialt og et økologisk domæne (se afsnit 3.2 og 6.10).
Det sociale domæne indeholder de aspekter af organismens funktion der refererer til dens overlevelse 'ud over sig selv'; de aspekter af organismens strukturelle koblinger der har med egen og fælles reproduktion at gøre, og dermed de dele af 'adfærden' der har at gøre med en samhørighed mellem organismer i forbindelse med en fælles overlevelse.
Det økologiske domæne indeholder de komplementære aspekter af organismens funktion; de aspekter af de strukturelle koblinger der har med individets egen overlevelse at gøre.
9.3 Fyletisk gradualisme eller ej:
Lad os i et tankeeksperiment se på en fylogenetisk række af organismer, hvor udsagn om organisationen af de sociale koblinger vedbliver at være sande i en vis periode; der sker altså ingen opsplitning i det sociale domæne, ingen reproduktiv artsdannelse.
Der kan i denne periode ske flere eller færre forandringer i udsagn om organisationen af de økologiske koblinger.
Sker der mange (eller 'store') forandringer vil vi (og det biologiske artsbegreb) have problemer med konflikten mellem slags og slægtskab.
For i et sådant tilfælde vil arten ikke udvise en stabil økologisk 'fremtræden' og/eller der vil ske skift i dens økologiske rolle (denne skelnen er Batesons skelnen mellem stabile relationer og stabile 'egenskaber' fra første afsnit).
At arterne på denne måde kan udvise gradvise ændringer i form og funktion uden at det ændrer artens samhørighed kaldes 'fyletisk gradualisme', og er en grundliggende opfattelse i darwinismen. Den hænger sammen med at naturlig udvælgelse anses for at være en stadigt forekommende og betydningsfuld proces i naturen.
Nye arter menes at opstå gennem en gradvis og kontinuert proces, styret af naturlig udvælgelse, hvor én art (slags) via en række mellemformer bliver til en ny. Kun hvis den gamle art overlever et eller andet sted (geografisk isolation), bliver der flere arter.
Sker der derimod få og små forandringer i organisationen af de økologiske koblinger hos organismerne i en socialt stabil periode, mens der sker flere i en periode med forandringer i organisationen af de sociale koblinger, vil slags og slægtskab være i større overensstemmelse; opsplitningen i det sociale domæne (de fylogenetiske forgreninger) falder i højere grad sammen med opsplitningen i slags. (Se endvidere afsnit 9.8)
Denne sidste opfattelse er grundlaget i teorien om 'punctuated equilibria' (afbrudte ligevægte).
9.4 Punctuated equilibria:
Teorien om afbrudte ligevægte har været central i evolutionsdebatten i det sidste årti. Tildels fordi den blev fremført i forbindelse med nogle grundliggende videnskabsteoretiske overvejelser; jeg har i opgaven berørt de fleste.
Teorien betragter arter som stabile enheder i evolutionen, og artsdannelsen som relativt kortvarige skift i arternes balance eller ligevægt.
Denne teori er ikke uforenelig med neodarwinismen.
Den betragter dog naturlig udvælgelse og arter i et perspektiv, der modsiger neodarwinismens fokus på populationsgenetikkens betydning i evolutionsteori.
Det neodarwinistiske fokus hænger sammen med Darwins analogi til avlsarbejde. Genetiske ændringer i en populationer under kunstig selektion beskrives særdeles godt af de populationsgenetiske modeller.
Neodarwinismen har (i ægte reduktionistisk ånd) betragtet sådanne ændringer i genhyppigheder i populationen som ansvarlig for alle evolutionsfænomener; og i forlængelse heraf det 'selviske gen' som den egentlige genstand for udvælgelse - intet højere (se også afsnit 9.10).
Der ligger flere spørgsmål i disse divergerende opfattelser af artsdannelsen: Dels spørgsmålet om forholdet mellem sociale og økologiske koblinger - om arternes stabilitet, og de mekanismer der kan ligge bag en sådan stabilitet; og dels spørgsmålet om forandringerne i det sociale domæne og de mekanismer der ligger bag den reproduktive opsplitning af arten i artsdannelsen.
Spørgsmålet om arternes faktiske stabilitet og stabilitetens tidshorisont er et område, hvor palæontologien (fossilerne og deres tolkning) er den eneste kilde til et svar. Vi kender alle eksemplerne på, at arter kan være stabile i en endog meget lang periode. Man har fundet såkaldte levende fossiler som Den Blå Fisk; og insekter, der er blevet bevaret i 30 mio. år indesluttet i rav, som kan henføres til en nulevende art.
Der er ikke enighed om den faktiske stabilitet endnu, man diskuterer heftigt fortolkningen af de palæontologiske data.
(Gould 77a, Gould 80 og Bonde 87, s 182ff)
9.5 Stabilitetens forklaringer:
Der er to grundliggende synsvinkler i en forklaring af stabilitet. En iboende forklaring som linedanserens homøostase; og en forklaring hvor begrundelsen ligger udenfor systemet, som når man balancerer en pind på fingeren.
Batesons grundliggende synspunkt er, at systemers i den levende verden opretholder deres stabilitet gennem begrænsninger i deres delsystemer (linedanseren), altså en iboende forklaring.
Der har været fremført mange forklaringer på artens stabilitet, på at arter kan være tilsyneladende uforandrede i millioner af år.
Den traditionelle populationsgenetiske forklaring er, at store populationer er stabile, fordi en stadig blanding af generne gennem panmiksi (at alle har mulighed for at parre sig med alle andre) forhindrer at tilfældige forandringer udbreder sig. Kun gener med en selektionsfordel vil brede sig i populationen. I et stabilt miljø vil arterne derfor være stabile.
Begge forudsætninger i forklaringen, panmiktiske populationer og stabile omgivelser har imidlertid kun en meget begrænset gyldighed.
Panmiktiske populationer er nok reglen i populationsgenetikken, men de er undtagelsen i naturen.
Men lad os se på det stabile miljø i stedet. Igen må vi klargøre hvilken tidshorisont vi bevæger os i. Befinder vi os i populationens tidshorisont, kan omgivelserne meget vel tænkes at være stabile. Men snakker vi om arter, og undersøger stabilitet i arternes tidshorisont, kan vi ikke snakke om stabile omgivelser. For her befinder sig jo netop artsdannelserne; medspillerne i det økologiske netværk er ikke konstante i denne tidshorisont (størrelsesordenen millioner af år).
Og da arternes stabilitet gælder både overfor tilpasninger og tilfældigheder, må den naturlige udvælgelse være sat mere eller mindre ud af spillet i den stabile periode.
Derimod kan relationerne eller de økologiske 'roller' være stabile udover artsdannelserne, som hestens fylogeni antyder. Som Bateson sagde ovenfor, er det jo netop gennem arternes ikke-stabilitet, arternes forandringer, at de økologiske relationer opretholdes.
At forklare artens stabilitet ved dens økologiske niche eller rolle lader sig således ikke gøre. Men argumentet ses ofte, fx skriver F.W. Brædstrup:
"Den fundamentale grund til arters faktiske konservatisme, må søges i det forhold, at de hver især har en bestemt niche, som de er indrettet til hårdnakket at bevare."
(Bonde 87, s 375)
Bevarelsen af nichen kan tværtimod være den fundamentale grund til arternes forandring!
Artsdannelser kan også indebære skift i de økologiske relationer, og en (i øvrigt stabil) art kan skifte økologisk rolle.
Dette kan ikke give os begrundelser for artens stabilitet, men er til gengæld centralt i en forståelse af artsdannelsen (se også afsnit 9.9).
At forklare artens konstans ved artens omgivelsers konstans er heller ikke gyldigt; arten og dens omgivelser er ikke et stabilt mekanisk system, men et dynamisk system. Der opstår masser af fluktuationer og forandringer i arten og dens omgivelser; stabiliteten begrænser disse fluktuationer, den er ikke en følge af manglende fluktuationer.
Der er også en anden side af de iboende forklaringer på stabilitet.
Asger Jorn generaliserede den norske antropolog Guttorm Gjessings udsagn om, at når en tradition omplantes til fremmede forhold, afsondret fra sin oprindelse, går udviklingen i stå; til også at omfatte biologiske forhold, han kalder det Gjessings regel:
"Enhver social organisme, der overlever sit oprindelige miljø og fortsætter sin eksistens ind i et nyt væsensforskelligt miljø, må være præget af en tendens til forstening, til at stivne i uforanderlig form."
Altså en forklaring på stabiliteten som et udslag af manglende interaktioner med et nyt miljø. Denne forklaring gælder formodentlig i visse tilfælde, men jeg tror den er en undtagelse. (Den modsvarende forklaring på forandring er meget nære koblinger til omgivelserne, se også afsnit 9.9)
(Heinberg i Bonde 87, s 199f)
Den iboende stabilitet må søges forklaret ved noget der ligner en indre homøostatisk mekanisme i arten.
Populationens overensstemmelse med omgivelserne kan ikke være årsagen til artens stabilitet på trods af forandringer i omgivelserne. Bonde skriver:
"Artens oplagte homøostasi skyldes således ikke genstrømmen, men snarere en indre homøostatisk effekt, knyttet til generne. Og det lyder rimeligt, for er der noget gener kan, så er det at kopiere. Hvordan de tillader det modsatte (evolution), ved vi stadig ikke rigtigt."
(Bonde 87, s 190)
Et eksempel på homøostatisk stabilitet af kvantitative egenskaber (egenskaber der ikke er udtryk for et enkelt gen) er fundet hos bananfluer. Kunstig selektion i en bestemt retning kan i mange generationer ske næsten uden forandringer; men så kan der pludselig ske en drastisk ændring over ret få generationer, efterfulgt af en ny periode med stabilitet. Dette eksempel giver jeg kun for at antyde, at organismens organisation som hele må være underlagt en endnu større konservatisme end sådanne enkelte egenskaber. Desuden viser eksemplet at ændringerne til gengæld kan ske pludseligt, når de sker. Organismens organisation er et netværk, hvor enkelte ændringer må modsvares af andre, hvis netværket skal fungere (se begrundelse herfor i afsnit 5.5).
(Parsons 83)
Cellens homøostase begrænser de genetiske mutationer og stabiliserer genomet; denne genetiske stabilitet er anerkendt og dens mekanismer er velkendte. Der opstår i massevis af mutationer og fejl i genomets kopiering, men i cellerne findes også en sammenligner, en 'reparations'mekanisme, der korrigerer ændringer i DNA-kædens nukleinsyresekvenser.
Men også på højere niveauer er der forslag til mulige mekanismer i homøostasens stabilitet.
Jeg omtalte i afsnit 5.3 en teori om meiosens betydning for komplekse organismer. I meiosen fjerner en lignende 'reparations'-mekanisme kønsceller, der har fået forstyrret den basale organisation i genomet eller indeholder for meget overflødig DNA.
Den hunlige organisme eller ægget lægger ligeledes en begrænsning på de hanlige kønsceller, fx sker der en kraftig selektion på planters pollen, når det vokser gennem blomstens støvfang og griffel.
En lignende mekanisme kan ligge til grund for spontane aborter af befrugtede æg eller fostre hos pattedyr.
Fælles for disse mekanismer er, at de kan betragtes som stokastiske processer, som jeg har defineret dem i afsnit 4.7. Og desuden, at den konvergente del af processen, sammenligneren eller den homøostatiske begrænsning, er meget effektiv. Dvs. udfaldet af de stokastiske processer er oftest meget forudsigeligt, i forhold til den underliggende divergente proces' uforudsigelighed.
Stokastiske processer kan altså opretholde en vis stabilitet som vi så det ovenfor, fordi de indeholder mekanismer der vælger det bekræftende. Men stokastiske processer er også potentielt nyskabende, da konvergensen ikke er fuldstændig.
9.7 Splittelse og social isolation:
Splittelse eller divergens er i sammenhæng med det ovennævnte det aspekt i en stokastisk proces, der er komplementært til stabiliteten. Nyskabelsen er den divergente proces' gennemtrængen udover den konvergente eller begrænsende proces. Splittelse er derfor ikke et fænomen, der er afgrænset til 'enheder' som arten, men et element i alle stokastiske processer på alle niveauer, og dermed i al kopiering og reproduktion. Splittelse eller nyskabelse kan derfor også foregå både i det økologiske og det sociale domæne, som det fremgik af tankeeksperimentet i afsnit 9.3.
Hvor splittelse finder sted i alle stokastiske processer, er isolation derimod en bestemt splittelse, der hører til i det sociale domæne. Og isolation kan kun bruges i denne mere begrænsede sammenhæng, i diskussionen af den specielle stokastiske proces der indgår i fælles reproduktion.
Isolation er i denne forbindelse et i høj grad relativt begreb.
Isolation må betegne enhver begrænsning i organismernes muligheder for at tage del i fælles reproduktion, lige fra mindre end total panmiksi til absolut geografisk eller tidslig isolation eller genetisk total uforenelighed (hvis der findes sådanne absolutter); altså i hele det sociale domænes udstrækning. Det sociale domæne er det område, der indeholder en balance mellem fælles reproduktion og isolation.
Problemet med et reproduktivt artsbegreb er tildels, at det må bygge på disse relative og komplementære begreber, isolation og reproduktivt fællesskab. Jeg har tidligere (i afsnit 3.2) set på, hvorfor reduktioner ikke kan give os svar på om der findes et potentielt reproduktivt fællesskab mellem to populationer.
Men udover dette eksperimentelle problem findes der også det problem, at et artsbegreb der bygger på fælles reproduktion langtfra er gyldigt for alle de manifestationer af det sociale domæne de levende væsener udviser.
Hvis vi laver en reductio ad absurdum, bliver problemet med det reproduktive artsbegreb mere tydeligt.
Vi kan se på nogle bakterier, der kun er i stand til at formere sig ved ukønnede delinger. Der kan godt opstå splittelse igennem disse delinger, bakteriernes fremtræden og økologiske rolle kan forandres.
I det sociale domæne er der på den ene side totalt reproduktivt fællesskab mellem bakterierne i en slægtskabslinie. En enkelt bakteries overlevelse og reproduktion er også reproduktionen af samtlige organismer i dens fylogenetiske fortid, tilbage til den første stammoder.
På den anden side har hver bakterie kun mulighed for at reproducere sig selv i fremtiden, isolationen set som fraværet af fremtidige fælles reproduktionsmuligheder er også total.
Det er indlysende at reproduktivt fællesskab med fylogenetiske forfædre ikke kan bruges som artskriterium, for så ender vi op med en eneste art, hvis det levende har én fælles afstamning. Og da muligheden for fremtidig fælles reproduktion er nul, er vores eneste anden mulighed at betragte hver bakterie som en selvstændig art.
Jamen, så har vi jo det økologiske kriterium, siger i.
Ja, men hvornår skal vi afgøre arten ud fra det sociale domæne, og hvornår ud fra det økologiske?
Bakterierne må nødvendigvis inddeles udfra deres økologiske fremtræden, men det var et ekstremt tilfælde og valget derfor let.
Hvad med alle de andre organismer, hvor det sociale domæne ikke har en så simpel struktur?
Vi kommer ikke uden om det centrale problem, - vores opdelinger er ikke 'naturlige' eller lovmæssige, vi må altid sige:
På den side så ..., men på den anden side ..., og på den tredje ...; og derefter blive enige om nogle foreløbige afgørelser, hvor det føles nødvendigt.
9.8 Opsplitning og isolationsbarrierer:
Jeg holder mig nu, som følge af ovennævnte overvejelser, til at diskutere organismer for hvilke et reproduktivt artsbegreb har en vis mening, og en diskussion af artsdannelsen som andet end fyletisk gradualisme derfor er mulig.
Jeg beskrev i afsnit 9.3 to muligheder for opsplitning (divergent udvikling) og isolation i artsdannelsen.
Dels en opfattelse af artsdannelsen som udløst af ydre faktorer, den geografiske isolation; og dels en indre baggrund, den reproduktive isolation. Det kan være vigtigt at skelne klart mellem den ydre og den indre isolation.
Der vil altid være nogen splittelse i arten, artens organismer er forskellige og de findes i en rumlig og tidslig organisation; der må derfor også være varierende grader af social isolation mellem artens organismer.
Den konventionelle opfattelse af artsdannelsen er den allopatriske ('et andet sted') model, der bygger på isolation gennem geografisk isolation. Hvis der senere tilfældigt opstår en indre isolation, kan de to arter leve i samme område uden at blandes, selvom den geografiske isolation forsvinder. Den naturlige udvælgelse kan da gennem konkurrence føre til en opsplitning af de to søsterarter, fx således at de deler den økologiske niche i området op imellem sig.
Eller de to arter kan vedblive at udfylde samme økologiske rolle i forskellige områder.
Modellen virker rimeligst for ikke alt for mobile organismer, der har mulighed for at blive rumligt isolerede i deres miljø. (Fx Bonde 87, s 169ff)
For nogle organismer og miljøer er den allopatriske model ikke rimelig. Claus Heinberg giver et eksempel med cichlider i Østafrikas store søer. Så godt som alle arterne findes kun i en enkelt sø; hovedparten af fiskene i søerne er cichlider; og søerne anslås kun at være 1-2 millioner år gamle. I hver sø findes der 100-200 cichlidearter i et komplekst økologisk netværk.
I et sådant tilfælde er en sympatrisk ('samme sted') model mere anvendelig. Heinberg foreslår en opsplitning pga. konkurrence mellem individerne, med indre isolation som en tilfældig følge. Nogle økologiske opsplitninger kan tænkes samtidigt at betyde rumlig isolation, fx ved opsplitning i forskellige habitater, og evt. efterfølges af en indre isolation.
På den anden side kan indre isolation (fx kromosom mutation eller ændring i social adfærd) også opstå først i en population, særligt hvis populationens sociale struktur betyder at organismerne ofte er (ydre) isoleret i mindre grupper i en generation eller mere, hvorved den indre isolation har mulighed for at udbrede sig til hele gruppen.
(Bonde 87, s 256ff og Gould 83b, s 337f)
Hvis vi ser på artsdannelse i et mere overordnet perspektiv, kan man se at artsdannelserne må ske i vidt forskellige situationer eller kontekster. Jeg tænker på tilstedeværelsen af og karakteren af allerede bestående økologiske relationer.
De østafrikanske søer må antages at have indeholdt et meget begrænset økologisk netværk i deres første levetid. Denne artsdannelseskontekst og de historiske tilfælde afspejles i de nuværende arter i søerne, der er 'udstrålet' fra en enkelt type.
Andre eksempler er Darwins finker på de vulkanske Galapagos-øer, og mange andre tilfælde hvor konteksten minder om situationen når en ny ø dukker op, og nye muligheder opstår.
I en længere tidshorisont finder vi lignende kontekster omkring livets opståen, tilblivelsen af organismer der kunne leve på landjorden, og andre lignende fundamentale nyskabelser i evolutionen; og ligeledes efter de sammenbrud i det globale økologiske netværk vi kender som masseuddøen.
I modsætning hertil må 'almindelig' artsdannelse i et komplekst netværk være afhængig af karakteren af de bestående økologiske relationer.
Konkurrence har længe været betragtet som den væsentligste økologiske interaktion; og konkurrencen mellem arter som en betydende mekanisme i den naturlige udvælgelse.
Konkurrence er en betegnelse for overlap i 'ligestillede' organismers økologiske relationer til 'lavere' niveauer (organismernes udnyttelse af ressourcerne), der resulterer i tilbagegang for begge parter eller fremgang for den ene part på den anden parts bekostning.
Mutualisme, dvs. overlap i økologiske relationer der giver fremgang for begge parter, anses traditionelt for at have langt mindre betydning i naturen.
Predation, her i en bred betydning som overlap i økologiske relationer til 'højere' niveauer (altså organismerne som ressourcer), er den komplementære mekanisme i de økologiske niveauers samspil. (Højere og lavere bruges her om forholdet mellem ressourcer og predatorer i de enkelte tilfælde; i det økologiske netværk som hele har højere og lavere ingen mening, hvis netværket er cirkulært organiseret.)
Det er således næppe meningsfuldt at diskutere konkurrence kontra predation i den naturlige udvælgelse som hele.
Derimod kan de økologiske relationer for de enkelte organismer og arter medvirke i en forståelse af deres tilblivelse.
Det er påvist, at to arter der udnytter de samme ressourcer kan blive mere forskellige, divergere eller splittes.
Deres karakteristika kan forskydes så konkurrencen mellem arterne formindskes.
Heinberg foreslog ovenfor at organismernes konkurrence indenfor arten på lignende måde kunne føre til opsplitning af arten.
Det forekommer rimeligt at en sådan opsplitning lettest kan foregå hvor mange alternative ressourcer endnu ligger uudnyttede hen, og konkurrence med andre arter ikke findes.
I mere komplekse og 'tætte' eller energieffektive økosystemer kan konkurrence derimod begrænse splittelse, idet nyskabelser oftest vil skulle bero på allerede udnyttede ressourcer. Konkurrencens 'funktion' kan altså udemærket skifte fra at være opsplittende til at være stabiliserende.
(Bonde 87, s 254ff og Schoener 82)
Man kan betragte økologiske nicher som potentielle muligheder for organismers eksistens (mulighed for at undgå hæmmende økologisk overlap), i de problemsituationer omkring overlevelse som livet består af.
Der ligger i denne opfattelse et svar på spørgsmålet om hvorfor evolutionen frembringer stadigt mere komplekse organismer.
Der findes niveauforskelle i de økologiske netværk og i organismernes organisation. Højere eller mere komplekse niveauer end de nuværende må ses som mulige men uudnyttede økologiske roller. Evolutionens 'retning' (med afbrydelser som masseuddøen) mod mere komplekse systemer, indenfor den begrænsning energistrømmen fra solen repræsenterer, må derfor tolkes på principielt samme måde som de simple dissipative strukturers udvikling, som tilfældighedernes udfoldelse.
I tilknytning til hvad jeg tidligere har sagt om kaos og tilfældighed, kan man se organismernes muligheder som det fremtidige modstykke til fortidens historiske valg af muligheder, bricolagen. Potentielle muligheder bliver til virkelige muligheder, - og andre mulige virkeliggørelser af den potentielle mulighed er ikke længere mulige. Livets opståen er fx ikke længere mulig i vores levende hav.
(Bl.a inspireret af Popper 81, s 136)
9.10 Stokastiske processer i forskellige tidshorisonter:
Jeg har i opgaven nævnt diskussionerne om hvilken enhed, der egentlig var evolutionsenheden, den enhed der var underlagt den naturlige udvælgelse.
Det er for tiden en udbredt opfattelse blandt neodarwinister, at genet er den egentlige genstand for naturlig udvælgelse; og man har forsøgt at forklare (hele!) evolutionen herudfra.
Richard Dawkins bog 'Det selviske gen' giver en populær fremstilling med genet som forklaringsgrundlag.
Der er dog også modstand mod denne ærkereduktionistiske opfattelse.
Ho og Saunders skriver blandt andet:
"Vi er enige med Dawkins i, at så længe man accepterer den neodarwinistiske ramme, er det selviske gen den logiske konklusion. Vi ser dog ikke dette som en triumf for teorien, men snarere som en reductio ad absurdum.
Neodarwinismen bygger på at der findes 'gener', skelnelige helheder som atomer, udsat for mutation og rekombination. Men hverken i strukturel eller funktionel forstand findes sådanne enheder. Dette må absolut betragtes som en fundamental svaghed ved teorien!"
(Oversat fra Ho 81, s 170)
Jeg håber at opgaven har belyst det meningsløse i at diskutere evolutionsenheder.
Jeg vil her argumentere for et mere righoldigt og forskelligartet syn på evolutionens mekanismer.
Naturlig udvælgelse blev af Darwin brugt om individernes indbyrdes konkurrence, og han brugte udtrykket seksuel udvælgelse for den tilsvarende mekanisme i det sociale domæne.
Der findes tilsvarende stokastiske processer på mange andre niveauer med forskellige tidshorisonter, men brugen af begrebet naturlig udvælgelse på alle disse niveauer fører efter min mening kun til en mudret diskussion.
Det er nemlig uhyre vigtigt at holde rede på de forskellige processers tidshorisont. Når jeg snakker om niveauer, inkluderer det en forskel i tids- eller rumhorisont for de indgående systemer.
Systemerne er netop karakteriseret ved deres tids/rumlige dynamik (se afsnit 7.3), og når man snakker om delsystemer må man også referere til de begrænsninger i tids/rum-horisont de udviser i forhold til systemet.
Eksistensen af stokastiske processer på lavere niveauer, specielt omkring mutationerne er som nævnt i afsnit 9.6 almindeligt accepteret. Også i det økologiske domæne findes velkendte stokastiske processer på lavere niveauer, og jeg har omtalt nogle som individets adaptation i afsnit 5.4.
Den naturlige udvælgelse har som nævnt også karakter af et fravalg; en begrænsning på individerne, der kun lader enkelte nyskabelser folde sig ud.
Der har derimod været megen uenighed og diskussion omkring eksistensen af stokastiske processer på højere niveauer end individets. Altså fremkomsten af højere systemer gennem divergente processer og begrænsningen heraf ved en sammenligning i en overlejrende konvergent proces. For fx kan genetikere nok acceptere at cellen udøver en begrænsning på de opståede mutationer, men ikke at fx økosystemet (eller noget tilsvarende) skulle udøve en lignede begrænsning på de opståede arterne.
Diskussionerne har specielt gået på artsudvælgelse, med en naturlig udvælgelse på tilfældige artsdannelser.
(Bl.a Hoffmeyer 80, 105ff)
Hvis vi skal finde sådanne processer må vi prøve at kigge efter kilder til nyskabelse og begrænsninger på denne nyskabelse.
Det kan være interessant her at se på det man traditionelt kalder underarter. Gould argumenterer for at underarter ikke er 'arter i svøb', men blindveje indenfor arten; og at artsdannelsen sker gennem andre mekanismer end dannelsen af underarter.
Men dannelsen af underarter er interessant i sig selv, som et eksempel på en 'højere' stokastisk proces.
Darwin gav mange eksempler på evolutionen af 'parringsstrukturer' gennem seksuel udvælgelse.
Nyere undersøgelser viser at den individuelle udvælgelse har betydning under udviklingen af disse sociale strukturer, men at der derefter indtræder en stabil periode, hvor individuel udvælgelse ikke influerer på strukturerne.
Robert Lacy omtaler et eksempel med monogame spurvefugle, hvor det har vist sig at arter uden kønsforskelle indeholder dobbelt så mange underarter som arter med farvestrålende hanner. Han diskuterer om disse fakta kan forklares udfra andet end en begrænsning på underarterne gennem en sammenligning af underarterne som grupper.
Sammenligningen kunne bero på de omkostninger for hannerne, der er forbundet med farvepragten.
(Lacy 85)
Jeg har tidligere omtalt den masseuddøen, der ret regelmæssigt har fundet sted på jorden. Disse perioder kan også ses som led i en uhyre stokastisk proces, hvor sammenligningen går på organismernes evne til at overleve drastiske forandringer i det jordiske miljø.
I denne sammenhæng dukker spørgsmålet om økosystemers diversitet (antal arter i forhold til antal organismer) og stabilitet op; masseuddøensperioderne må betragtes som et sammenbrud af de globale økosystemers stabilitet.
Det er velkendt, at der i troperne findes økosystemer med en ofte langt højere diversitet end i de koldere klimater.
Her kan det være givtigt at tænke på Maturanas opfattelse af individernes overensstemmelse med omgivelserne; den sammenlignende proces i den naturlige drift.
Denne (for simple) analogi kan lede til et fokus på økosystemets overensstemmelse med dets 'omgivelser'.
Omgivelsernes stabilitet er en afgørende faktor i homøostasens regulering, stabile ydre forhold fører til en mere grundliggende, og derfor mere 'økonomisk' og mere rigid regulering i systemet.
Økosystemers diversitet hænger sammen med arternes økologiske fleksibilitet, højdiverse systemer består af specialiserede arter, mens lavdiverse systemer indeholder flere generalister. Regnskovenes stabilitet er ikke en stabilitet over for miljøforandringer, men netop en historisk stabilitet af miljøets betingelser.
Gennem denne stabile historie har økosystemet udviklet sig til et meget økonomisk og ufleksibelt system, med en høj udnyttelse af den potentielle strøm af information, herunder energi, gennem systemet; der er udviklet et netværk af mange arter og sket en specialisering af de enkelte arters funktion.
Andre økosystemer eksisterer i mere variable sammenhænge, systemerne er ikke helt så økonomiske men til gengæld er de meget fleksible overfor forandringer i miljøet, fordi arterne er færre og hver især har en mere generel funktion.
Jeg vil ikke komme med flere eksempler på større stokastiske processer her, men i stedet se lidt på de enkelte levende væseners tidshorisonter.
Det er nemlig også væsentligt, i forbindelsen med omtalen af stokastiske processer i forskellige tidshorisonter, at gøre opmærksom på at levende væsener lever i meget forskellige tidshorisonter.
Vi mennesker lever, som de andre levende væsener der er bundet til den kønnede formering, med den begrænsning for vores individuelle livsforløb der hedder aldring.
Men der findes andre levende væsener der ikke lever under denne begrænsning.
En stor del af klodens organismer reproducerer sig mest eller udelukkende ukønnet gennem mitoser; og lever derfor i en tidshorisont, og en verden af potentiel ubegrænsethed, der er ufattelig meget anderledes end vores egen.
(Se også Valen 87 og Cook 86)
De fleste forklaringer og det meste af vores forståelse af evolutionen refererer til kønnede organismer. Denne opgave adskiller sig ikke meget i denne retning, for det er en langvarig proces at forstå de aspekter af det levende der er radikalt forskellige fra os selv.
AFSLUTTENDE BEMÆRKNINGER Indhold
Jeg kan bedst beskrive min personlige oplevelse af det at skrive denne opgave, som en slags 'Columbusæg'.
Ikke i den forstand at ideen skulle være mere eller mindre unik; men jeg genkender de følelser der opstår før og efter ægget er blevet stillet på enden.
I starten var opgaven og emnerne helt uoverskuelige, og problemerne tilsyneladende umulige at løse.
Nu da opgaven er færdig, ser mange af problemerne næsten banale ud; og de enkelte dele af opgaven virker omtrent som gentagelser af, hvad kloge mennesker forlængst har tænkt.
På den anden side er der stadig lige mange og mindst lige så uoverskuelige problemer tilbage; men jeg føler samtidigt at mine muligheder for og min lyst til at tage fat på disse problemer er større end de var før.
Jeg vil slutte af med et uddrag af en tale Niels Bohr holdt ved studenterjubilæet den 21. september 1928.
Uddraget udtrykker en stor optimisme på trods af eller netop på grund af, at selv nok så store opgavers løsning fører os tilbage til et sted, der i hvert fald minder stærkt om det sted hvor vi begyndte.
Bohrs tanker om harmoni klinger også sammen med en 'etik', der længe har ligget i mine følelser og tanker omkring den retning vores verdens udvikling tager.
Jeg tror på, at ødelæggelse af harmonien er forkert; og derfor grunder min tro på hvad vi bør gøre, på en respekt for det der er og har været. Jeg tror ikke vi kan leve uden den harmoni vi er oprundet af.
Fra Niels Bohrs tale ved studenterjubilæet:
"Det er ikke erkendelsen af vor menneskelige begrænsning, men bestræbelserne på at udforske denne begrænsnings natur, der præger vor tid. Det vil kun give os en fattig forestilling om vore muligheder, om vi ville sammenligne vor begrænsning med en uoverstigelig mur, der på bestemte veje er sat for vore tanker.
Overalt åbner sig nye udblik, og nye sammenhæng lader sig erkende, men enhver vej, som vi slår ind på, deler sig atter og atter og krummer sig, så at vi hurtigt taber retningen og før eller senere kommer tilbage til vort udgangspunkt.
Alligevel kan vi altid vende hjem med udbytte, og for rigdommen af det indhold, som vi kan samle og ordne, ser vi ingen grænser. Ud fra dybere og dybere udforskede forudsætninger erkendes større og større sammenhæng.
Således opfattet lever vi under et stadig rigere indtryk af en evig og uendelig harmoni; vel at forstå, harmonien selv lader sig kun ane, men aldrig gribe; ved ethvert forsøg derpå svinder den efter sit væsens natur ud af vore hænder.
Intet er fast, hver tanke, ja selv hvert ord er kun egnet til understrege en sammenhæng, der i sig selv aldrig kan fuldt beskrives, men altid uddybes."
(Bohr 85, s 262f)
ORDLISTE Indhold
I ordlisten forekommer fremmedord og begreber der er specielle for de emneområder jeg har været inde på i opgaven. Ofte vil ordet eller begrebet være forklaret første gang det optræder i teksten, og jeg vil da blot henvise hertil; kun i undtagelserne herfra findes en forklaring her i ordlisten.
a priori: Uafhængig af eller forud for erfaringen (a priori syntetiske udsagn: Udsagn om virkeligheden der er uafhængige af erfaringen.)
autopoiese 6.2
analog kommunikation 4.3
determinisme: Troen på at alting er forudbestemt, at det følger direkte af det forudgående.
digital kommunikation 4.3
divergent proces 4.7
DNA: Fra engelsk - Deoxyribo Nucleic Acid ('ribose-kernesyre'), det stof der udgør arvematerialet.
ekstrapolation 3.5
emergens: Skabelse, opdukken, noget nyt bliver til.
emisk aspekt 2.2
empirisk: Udfra erfaringen.
epigenese 5.1
epistemologi: Erkendelseslære, læren om grundlaget for viden.
eukaryoter: De levende organismer inddeles i to hovedgrupper; prokaryoter var de første levende celler, senere opstod eukaryoterne, formodentlig af kolonilevende eller flercellede prokaryoter. Prokaryoter er oftest mindre end eukaryoter, de har et ringformet genom og ingen 'rigtige' organeller. Eukaryoter har en cellekerne med et genom der er opdelt i ikke-ringformede kromosomer, og de har forskellige andre (prokaryot-lignende) organeller som mitochondrier og grønkorn.
falsifikation 1.4
fænotype 4.5
fylogenese 3.4
genom: Cellens eller organismens 'sæt' af DNA.
genotype 4.5
hierarkisk 4.1
homøostase 5.1
ikke-viden 1.8
induktion 1.4
interaktion: Vekselvirken eller gensidig påvirkning.
intrikate: Indviklede og delikate.
komplementaritet 2.6
kontekst 4.4
konvergent proces 4.7
kybernetik: Videnskaben om styringsprocesser, af det græske ?'kybernetes' der betyder styrmand.
logisk type 4.2
matrice 2.9
meiose 5.2
mitose 5.2
morfologi: Læren om organismernes form og bygning.
naturalisme: Her - den opfattelse at naturen kan fortælle os noget om hvad mennesker bør og ikke bør.
nukleotider: Et led i nukleinsyre-kæden; består af en kvælstofholdig base, ribose (i RNA) eller deoxyribose i DNA), og et molekyle fosfat.
ontogenese 3.4
ontologi: Læren om det værende, om det der eksisterer.
palæontologi: Videnskaben om fossiler og fortidige organismer.
paradigme 1.3
perception: Sansning eller opfattelse.
prokaryoter: Se eukaryoter.
reductio ad absurdum: Tilbageføring (via logiske slutninger) til ?det meningsløse.
reduktionisme 1.8
reproduktion 5.2
RNA: Overfører informationer fra DNA til ribosomerne, hvor de ?oversættes til proteinernes aminosyresekvenser.
somatisk 5.1
system 4.1
stokastisk proces 4.7
tautologi 2.9
tidshorisont 3.5
translation 4.5
transskription 4.5
verifikation 1.4
LITTERATURLISTE Indhold
Jeg vil gerne her anbefale nogle bøger, som jeg især har fået et stort udbytte af at læse.
Som det fremgår af opgaven har Gregory Bateson inspireret mig meget, og jeg føler at jeg kan blive ved med at hente inspiration og forståelse ud af hans bøger; jeg har også medtaget den nyeste, som hans datter har udgivet efter hans død, og som jeg for nylig har fået. Ølgaards bog giver en god og dybtgående gennemgang af både Ånd og Natur og den ikke-oversatte artikelsamling Steps to an Ecology of Mind på dansk.
Desuden kan jeg anbefale Maturanas og Hoffmeyers bøger, der i deres indhold berører mange af de emner jeg har været inde på i denne opgave.
Akimoto, Shin-ichi 1985:
Occurrence of abnormal phenotypes in a host-alternating aphid and their implications for genome organisation and evolution.
- Evolutionary Theory vol. 7, 179-194. Chicago.
Ashby, W. Rose 1979:
An introduction to Cybernetics.
Methuen & Co LTD, London
Bateson, Gregory 1984:
Ånd og Natur.
Rosinante.
Bateson, Gregory 1985:
Steps to an ecology of mind.
Ballantine Books, New York.
Bateson, Gregory and Mary Catherine Bateson 1988:
Angels Fear. Towards an epistemology of the sacred.
Bantam Books.
Bertalanffy, Ludwig von 1975:
Perspectives on General System Theory.
George Braziller. New York.
Bohm, David 1986:
Helhed og den indfoldede orden.
Forlaget ASK.
Bohr, Niels 1985:
Naturbeskrivelse og menneskelig erkendelse.
Rhodos.
Bonde, Niels og Henrik Stangerup (eds) 1985:
Naturens historiefortællere I. Udviklingsideens historie. Fra Platon til Darwin.
G E C Gads Forlag.
Bonde, Niels og Jesper Hoffmeyer (eds) 1987:
Naturens historiefortællere II. Udviklingsideens historie. Fra Darwins syntese til nutidens krise.
G E C Gads Forlag.
Cook, Robert E. 1986:
Population biology and evolution of clonal organisms.
Yale University Press. New Haven.
Cracraft, Joel 1983:
Cladistic analysis an vicariance biogeography.
- American Scientist vol. 71, 273-281.
Dammann, Erik 1987:
Bak tid og rom.
Dreyers Forlag A/S, Oslo.
Darwin, Charles 1979:
The illustrated Origin of Species.
Faber and Faber, London.
Davies, Paul 1987:
Gud och den nya fysiken.
Bokforlaget Prisma. Stockholm.
Dawkins, Richard 1977:
Det selviske gen.
Fremad.
Dobzansky, Theodosius, Fransisco Ayala, Ledyard Stebbins and James Valentine 1977:
Evolution.
W H Freeman and Company, San Fransisco.
Ettinger, Lia 1986:
Meiosis: A selection stage preserving the genomes pattern of organization.
- Evolutionary Theory vol. 8, 17-26. Chicago.
Fjerdingstad, E.J. 1988:
Et tilfælde af teknofobi.
Kronik i Politiken 10/3 88.
Gamlin, Linda og Gail Vines (eds.) 1987:
Videnskabens Verden. Den levende verden.
Politikens Forlag.
Gould, S. J. and Niles Eldredge 1977a:
Punctuated equilibria: The length and mode of evolution reconsidered.
- Paleobiologi vol. 3, 115-151. Chicago.
Gould, Stephen Jay 1977b:
Ontogeny and phylogeny.
The Belknap Press of Harvard University Press, Cambridge, Mass.
Gould, Stephen Jay 1978:
Ever since Darwin. Reflections in natural history.
Burnett Books Limited, London.
Gould, Stephen Jay 1980:
Is a new and general theory of evolution emerging?
- Paleobiology vol. 6, 119-130. Chicago
Gould, Stephen Jay 1983a:
The pandas thumb. More reflections...
Penguin Books.
Gould, Stephen Jay 1983b:
Hens Teeth and Horses Toes.
W. W. Norton & Co., New York.
Gould, Stephen Jay 1985:
The Flamingos smile. Reflections in ....
W. W. Norton & Co., New York.
Ho, Mae-Wan and Peter T. Saunders 1981:
What is the unit of natural selection?
- Evolutionary Theory vol. 5, 169-172. Chicago.
Hoffmeyer, Jesper 1980:
Evolution, økologi, historie. Neodarwinismens krise.
politisk revy.
Hoffmeyer, Jesper 1984:
Naturen i hovedet.
Rosinante.
Hoffmeyer, Jesper 1988:
DNA er ikke Gud.
Kronik i Politiken 25/2 88.
Jacob, Francois 1985:
Mulighedernes spil. Om det levendes mangfoldighed.
Hekla.
Jantsch, Erich 1980:
The self-organizing universe.
Pergamon Press.
Jørgensen, Jørgen 1978:
Havebrugszoologi a.
DSR Forlag, København.
Kragh, Helge og Stig Andur Pedersen 1981:
Naturvidenskabsteori.
Nyt Nordisk Forlag Arnold Busck A/S.
Kuhn, Thomas 1973:
Videnskabens revolutioner.
Fremad.
Lacy, Robert C. 1985:
Evidence that group selection counters the evolution of sexual dimorphism.
- Evolutionary Theory vol. 7, 173-177.Chicago.
Lovelock, James 1987:
Gaia - Et nyt syn på Jordens liv.
Hovedland.
Lowenstein, Jerold M. 1985:
Molecular approaches to the identification of species.
- American Scientist vol. 73, 541-547.
Magee, Bryan 1973:
Karl Popper.
Nyt Nordisk Forlag Arnold Busck A/S.
Maturana, Humberto og Fransisco Varela 1987:
Kundskabens træ. Den menneskelige erkendelses biologiske rødder.
Forlaget ASK.
Mayr, Ernst 1982:
The growth of biological thought.
The Belknap Press of Harvard University Press, Cambridge, Massachusetts.
Monod, Jacques 1971:
Tilfældigheden og nødvendigheden.
Fremad.
Naylor, Bruce G. and Paul Hanford 1985:
In defence of Darwins theory.
- Bioscience vol. 35, no. 8.
Nørretranders, Tor 1985:
Det udelelige.
Gyldendal.
Nørretranders, Tor 1987:
Naturvidenskab og ikke-viden.
Kimære.
Parsons, Peter A. 1983:
The genetic basis of quantitative traits: Evidence for punctuational evolutionary transitions at the intraspecific level.
- Evolutionary Theory vol. 6, 175-184. Chicago.
Pirsig, Robert M. 1985:
Zen og kunsten at vedligeholde en motorcykel.
Borgen.
Popper, Karl R. 1973:
Objective Knowledge. An evolutionary approach.
Oxford University Press, Oxford.
Popper, Karl R. 1981:
Fornuft og rimelighet som tenkemåte. Utvalgte essays.
Dreyers Forlag, Oslo.
Prigogine, Ilaya og Isabelle Stengers 1985:
Den nye pagt mellem mennesket og universet. Nye veje i naturvidenskaberne.
Forlaget ASK.
Ravn, Ib 1988:
Fra natur til kultur.
Kronik i Politiken 12/10-88.
Schoener, Thomas W 1982:
The controversy over interspecific competition.
- American Scientist vol. 70, 586-591.
Sheldrake, A. Rupert 1981:
The hypothesis of formative causation.
Bond & Briggs, London.
Snyder, L.A., D. Freifelder and D.L. Hartl 1985:
General genetics.
Jones and Bartlett Publishers, Inc., Boston.
Thom, René 1975:
Structural stability and Morphogenesis.
Benjamin. Reading, Mass.
Valen, Leigh M. Van 1987:
Non-Weismannian Evolution.
- Evolutionary Theory vol 8, 101-107. Chicago.
Whitehead, A.N. and Bertrand Russel 1910-13:
Principia Mathematica.
Cambridge University Press, Cambridge.
Zinkernagel, Peter 1957:
Omverdensproblemet.
G.E.C. Gads Forlag.
Ølgaard, Bent 1986:
Kommunikation og økomentale systemer – ifølge Gregory Bateson.
Forlaget ASK.